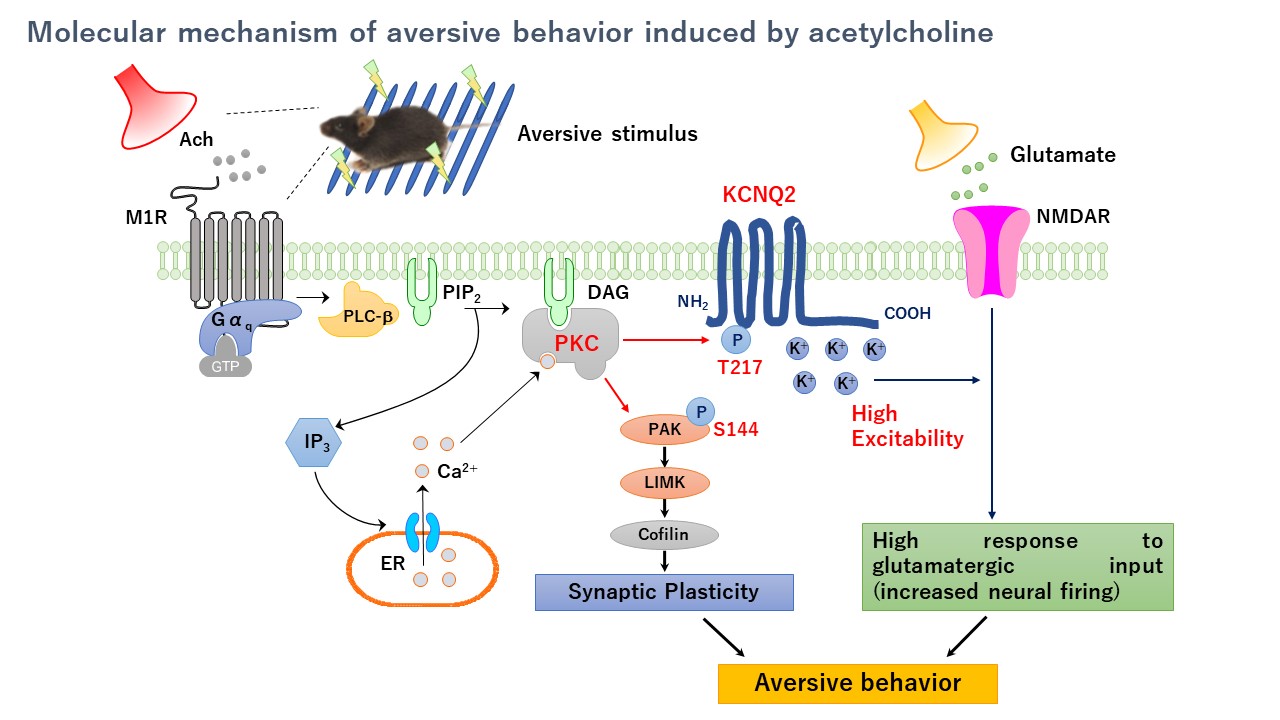
Как BK-каналы волосковых клеток взаимодействуют с белком RACK1. Какое влияние оказывает протеинкиназа C на экспрессию BK-каналов на поверхности клетки. Какие механизмы лежат в основе этих процессов. Почему это важно для функционирования волосковых клеток внутреннего уха.
Содержание
BK-каналы и их роль в функционировании волосковых клеток
BK-каналы (каналы большой проводимости, активируемые кальцием и напряжением) играют важную роль в функционировании волосковых клеток внутреннего уха. Эти каналы участвуют в:
- Настройке частотной избирательности волосковых клеток
- Формировании электрического резонанса
- Регуляции высвобождения нейромедиатора
Поэтому регуляция активности и поверхностной экспрессии BK-каналов имеет большое значение для слуховой функции.
Взаимодействие BK-каналов с белком RACK1
В ходе исследований было обнаружено, что BK-каналы волосковых клеток взаимодействуют с белком RACK1 (рецептор активированной С-киназы 1). Это взаимодействие имеет важное функциональное значение:
- RACK1 служит в качестве каркасного белка, обеспечивая правильную локализацию BK-каналов в клетке
- Взаимодействие с RACK1 способствует стабилизации BK-каналов на поверхности клетки
- RACK1 может регулировать активность BK-каналов, влияя на их фосфорилирование
Методы изучения взаимодействия BK-каналов и RACK1
Для изучения взаимодействия BK-каналов и RACK1 исследователи использовали следующие методы:
- Коиммунопреципитация для выявления белок-белковых взаимодействий
- Флуоресцентная микроскопия для визуализации колокализации белков
- Электрофизиологические методы для оценки функциональных эффектов
Эти методы позволили подтвердить прямое взаимодействие BK-каналов и RACK1 в волосковых клетках.
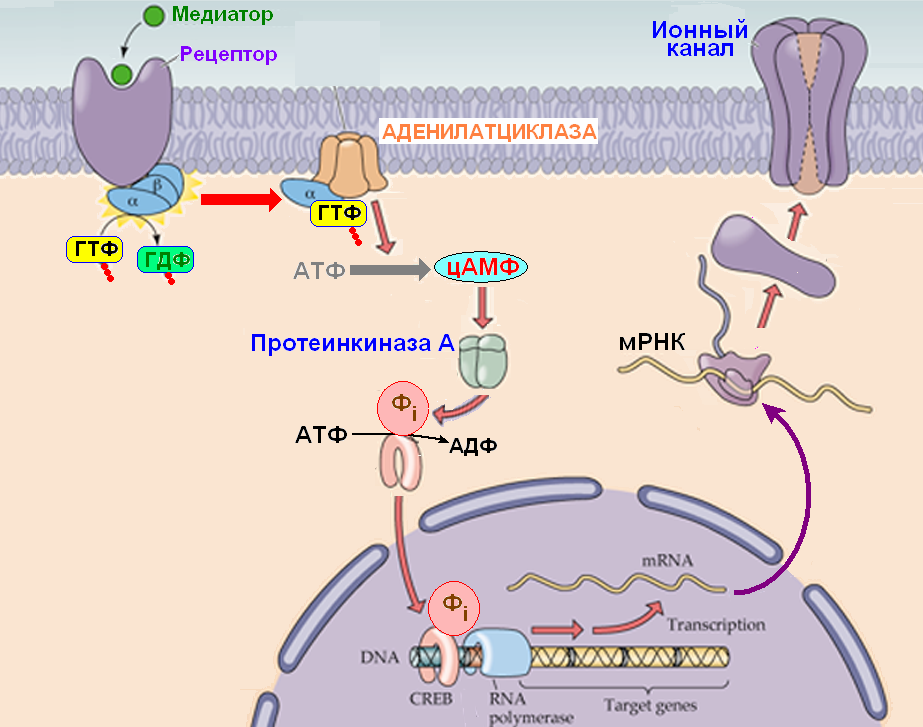
Влияние протеинкиназы С на экспрессию BK-каналов
Исследования показали, что протеинкиназа С (PKC) оказывает значительное влияние на поверхностную экспрессию BK-каналов в волосковых клетках:
- Активация PKC приводит к увеличению количества BK-каналов на поверхности клетки
- Этот эффект опосредован непрямым фосфорилированием BK-каналов
- PKC фосфорилирует другие белки, которые затем влияют на трафик BK-каналов к мембране
Механизмы влияния PKC на экспрессию BK-каналов
Выявлены следующие механизмы, через которые PKC регулирует поверхностную экспрессию BK-каналов:
- Фосфорилирование белков цитоскелета, участвующих в транспорте везикул с BK-каналами
- Модификация белков эндоцитоза, что замедляет интернализацию каналов
- Активация транскрипционных факторов, усиливающих экспрессию BK-каналов
Таким образом, PKC оказывает комплексное влияние на различные этапы жизненного цикла BK-каналов в клетке.
Физиологическое значение регуляции BK-каналов
Регуляция активности и поверхностной экспрессии BK-каналов имеет важное физиологическое значение для функционирования волосковых клеток:
- Обеспечивает тонкую настройку частотной избирательности
- Позволяет адаптировать чувствительность клеток к изменяющимся условиям
- Участвует в процессах пластичности слуховой системы
Нарушения в регуляции BK-каналов могут приводить к различным слуховым расстройствам.
Роль BK-каналов в патологии слуха
Исследования показали, что дисфункция BK-каналов может быть связана с рядом патологических состояний:
- Нейросенсорная тугоухость
- Шум в ушах (тиннитус)
- Нарушения равновесия
Поэтому понимание механизмов регуляции BK-каналов важно для разработки новых методов лечения слуховых расстройств.

Перспективы дальнейших исследований
Дальнейшие исследования взаимодействия BK-каналов с RACK1 и регуляции их экспрессии протеинкиназой C могут быть направлены на:
- Выявление конкретных сайтов взаимодействия BK-каналов и RACK1
- Идентификацию всех белков-мишеней PKC, влияющих на BK-каналы
- Изучение роли этих механизмов в различных типах волосковых клеток
- Поиск способов фармакологической модуляции активности BK-каналов
Эти исследования могут открыть новые возможности для коррекции нарушений слуха и вестибулярной функции.
Методы исследования BK-каналов в волосковых клетках
Изучение BK-каналов в волосковых клетках представляет определенные сложности из-за малых размеров этих клеток и их труднодоступности. Однако разработан ряд эффективных методов исследования:
Электрофизиологические методы
- Метод локальной фиксации потенциала (patch-clamp) позволяет регистрировать токи через отдельные BK-каналы
- Внутриклеточная регистрация позволяет изучать вклад BK-каналов в формирование потенциала действия волосковых клеток
Молекулярно-биологические методы
- РНК-интерференция для подавления экспрессии BK-каналов
- Создание трансгенных животных с модифицированными BK-каналами
- Количественная ПЦР для оценки уровня экспрессии генов BK-каналов
Методы визуализации
- Иммуногистохимическое окрашивание для выявления локализации BK-каналов в клетке
- Конфокальная микроскопия для изучения динамики BK-каналов в живых клетках
- Электронная микроскопия для исследования ультраструктуры комплексов BK-каналов
Комбинация этих методов позволяет получить всестороннее представление о функционировании и регуляции BK-каналов в волосковых клетках.
Эволюция BK-каналов в волосковых клетках
BK-каналы играют важную роль в функционировании волосковых клеток у различных видов животных. Изучение эволюции этих каналов позволяет лучше понять их значение:
- BK-каналы обнаружены в волосковых клетках всех позвоночных, от рыб до млекопитающих
- У низших позвоночных BK-каналы участвуют преимущественно в детекции вибраций и гравитации
- У высших позвоночных BK-каналы приобрели дополнительные функции в обработке звуковых сигналов
- Появление альтернативного сплайсинга генов BK-каналов у млекопитающих обеспечило более тонкую настройку их свойств
Эволюционный подход помогает понять, почему BK-каналы столь важны для слуховой функции у человека и других млекопитающих.
Заключение
Исследования взаимодействия BK-каналов с белком RACK1 и влияния протеинкиназы C на их экспрессию открывают новые аспекты функционирования волосковых клеток внутреннего уха. Эти механизмы обеспечивают тонкую регуляцию активности BK-каналов, что критически важно для нормальной слуховой функции. Дальнейшее изучение этих процессов может привести к разработке новых методов диагностики и лечения различных нарушений слуха.

Хомяк Радде, как житель дикой степи!
У хомяка Радде есть еще несколько названий – предкавказский и дагестанский. Ближайшим родственником-братом ему является сирийский хомяк. Но от него Радде отличается гораздо более крупным размером: от 170 до 280 мм в длину.
Уже по дополнительным названиям понятно, что распространен он в горной местности, а, как правило, обитатели гор, в сравнении с равнинными жителями, более крупных размеров.
В окрасе хомяка Радде преобладает коричневый цвет с присутствием темных и светлых отметин. Брюшко черненькое, за ушами и на щеках шерстка слегка высветлена.
Места обитания
Хомяк Радде – степной житель. Степи – это изобилие различных многолетних трав, что очень привлекательно для него. Любит селиться он на землях, засеянных злаковыми культурами. Зачастую и нору он строит вблизи распаханного поля. Ему пригодны для жизни и лесополосы, но только территории, не подвергающиеся затоплению.
Образ жизни и питание
Как и все хомяки, Радде ведет преимущественно ночной образ жизни. Дневной свет его не радует, и на поверхность днем он выходит крайне редко, и только в теплое время года. Норы у этого вида хомяков отличаются глубиной. Горные разновидности Радде снабжают их несколькими выходами, а равнинным достаточно одного, через который они и выходят на поверхность. Норы роются с разнообразными ответвлениями, которые начинаются на метровой глубине. Всех хомяков, и Радде не исключение, отличает редкая хозяйственность. Они очень запасливы и хранят в своих кладовых до 16 кг разнообразного корма. Так что к выходу из спячки Радде готовятся со всей ответственностью и при выходе из нее заготовленный провиант сразу же съедается.
В летний период хомяк питается травами и зелеными растениями. Горох, клевер, горная астра, люцерн и т. д. охотно поедаются им. Зерна пшеницы или ячменя тоже вполне пригодны для него. К концу лета созревают бобовые, различные корнеплоды, и Радде переключается на них. Конец лета – начало осени — период накопления запасов на зиму, когда он впадет в спячку. Проспать хомяк может от 4 до 6 месяцев – все зависит от того, насколько долго продержится холодная погода.
Конец лета – начало осени — период накопления запасов на зиму, когда он впадет в спячку. Проспать хомяк может от 4 до 6 месяцев – все зависит от того, насколько долго продержится холодная погода.
Размножение и потомство
Хомяков отличает редкая плодовитость. Потомство приносят они несколько раз в год: равнинные разновидности – до 4 раз, горные – дважды. Помет от одной самки может насчитывать в среднем 20 детенышей. Так что природа хорошо подстраховала хомяков от вымирания. Вряд ли оно когда-нибудь будет им угрожать при такой плодовитости.
Враг сельского хозяйства и переносчик инфекций
Последствия деятельности Радде, предпочитающего селиться вблизи сельхозугодий, опустошительны. В 50 кв. м от его норы растительность практически отсутствует. Так что земледельцы такому соседству совсем не рады: хомяк приносит им серьезные убытки.
Но и это еще не все. Хомяк Радде еще и переносчик инфекции. Он заражает туляремией не только животных, но и человека. Так что избавиться от него стремятся и по этой причине. Для этого используют различные ловушки, заливают водой норы, просто отлавливают хомяков ради его шкурки, которую используют для изготовления одежды.
Так что избавиться от него стремятся и по этой причине. Для этого используют различные ловушки, заливают водой норы, просто отлавливают хомяков ради его шкурки, которую используют для изготовления одежды.
- < Назад
- Вперёд >
Хомяк Радде
Оглавление
- Особенности внешнего вида
- Поведенческие инстинкты
- Образ жизни
- Питание
- Размножение
- Вред для сельскохозяйственных угодий
Более двухсот видов включает в себя семейство Хомяковых. Наряду с известными домашними хомячками в природе существуют дикие особи. Одним из таких является хомяк Радде. Относится к виду Средних хомяков. Своё научное наименование получил по имени учёного-открывателя Г.И. Радде. В природе обитает в Грузии, России, Кавказе, горных степях Дагестана, Предкавказье. Ландшафт выбирает степной, предгорный, лесостепной. Селится на обжитых человеком территориях, поедая урожай в огородах, на дачах, полях. В народе грызуна прозвали диким хомяком за его крутой нрав и агрессивные повадки. Примечательно, что на этом наименования зверька не заканчиваются. На территории Кавказа его именуют – Предкавказский хомяк, по основному месту его распространения. Жители предгорных районов Дагестана называют его хомяк Дагестанский. Множественные наименования оправданы и обозначают один вид грызуна, о котором пойдёт речь в статье.
В народе грызуна прозвали диким хомяком за его крутой нрав и агрессивные повадки. Примечательно, что на этом наименования зверька не заканчиваются. На территории Кавказа его именуют – Предкавказский хомяк, по основному месту его распространения. Жители предгорных районов Дагестана называют его хомяк Дагестанский. Множественные наименования оправданы и обозначают один вид грызуна, о котором пойдёт речь в статье.
Особенности внешнего вида
Длина тела грызуна достигает 28 см, хвоста 14 мм. Имеет небольшие круглой формы уши, подошва лап полностью голая, хвост покрыт длинными грубыми волосами. Спина может быть рыжего, либо бурого цвета, грудь и брюшко чёрные. От уха и до плеча идёт длинное чёрное пятно, в середину которого вклинивается небольшое пятнышко молочного цвета. Такие же молочные пятна имеются на щеках и боках зверька. Лапки в основном белого цвета. В природе встречаются полностью чёрные особи или альбиносы. Предкавказский хомяк – обладатель очень красивой и качественной шкурки. Её мех густой, блестящий, шелковистый. Как и другие грызуны этого вида имеет защечные мешочки, с помощью которых переносит корм.
Её мех густой, блестящий, шелковистый. Как и другие грызуны этого вида имеет защечные мешочки, с помощью которых переносит корм.
Поведенческие инстинкты
Хомяк Радде обладает плохим зрением, но отличным слухом и нюхом. Отличается агрессивным нравом и представляет угрозу для человека и домашних животных, справиться с ним может только собака. Попытки человека приручить его всегда заканчивались провалом.
Грызун обладает неимоверной живучестью. Не боится морозов, так как имеет густой подшерсток. При малейшей опасности старается укрыться в норе, но если такое невозможно, смело бросается на врага. Встаёт на задние лапки, передние поднимает вверх и выпускает когти, при этом делает страшный оскал. Может кидаться и подпрыгивать. Жертву или обидчика кусает, оставляя на теле рваные ранки. Для человека хомяк Радде опасен тем, что является переносчиком тяжёлой инфекции под названием туляремия, которая характеризуется поражением лимфатических узлов, лихорадкой и интоксикацией организма.
Образ жизни
Это сумрачное животное. Ведёт ночной образ жизни, днём увидеть его практически невозможно. Роет глубокие витиеватые норы, с множеством выходов и отсеков. Очень хозяйственный, каждый отсек в норке имеет своё предназначение – для проживания, для хранения запасов, отхожее место. Грызун никогда не занимает чужие норы, предпочитая рыть их самостоятельно, умело маскируя ходы и выходы. Зимой впадает в спячку, но периодически просыпается, чтобы насытить организм. Спячка длится примерно шесть месяцев в зависимости от состояния погоды.
В природных условиях могут прожить до пяти лет, что случается редко, ведь у дикого хомяка очень много врагов:
- Лисы;
- Совы;
- Канюки;
- Соколы;
- Коршуны.
Грызуну присущ одиночный образ жизни. Не терпит присутствия сородичей и активно борется за свою территорию. Только во время спаривания грызуны могут проживать в одной норке, но это вызвано необходимостью, как только она отпадает, они расходятся по своим жилищам. Интересно, что в этот период самец защищает территорию самки, никому не позволяя заселиться во временно опустевшую нору. Самцы полигамны, одновременно могут спариваться с несколькими самками.
Интересно, что в этот период самец защищает территорию самки, никому не позволяя заселиться во временно опустевшую нору. Самцы полигамны, одновременно могут спариваться с несколькими самками.
Питание
Предкавказский хомяк всеяден. Уничтожает всё съестное на территории до 50 квадратных метров от своего жилища. Является любителем кормовых культур, выращиваемых человеком для корма домашнего скота. К ним относят:
- Силосные культуры – кукуруза, подсолнечник.
- Многолетние травы, выращиваемые для получения сена, силоса, летних пастбищ скота.
- Корневые культуры – свёкла, морковь, брюква.
- Бахчевые культуры — дыни, арбузы, тыква, кабачок.
На зиму хомяк заготавливает зерновые культуры: пшеницу, ячмень, рожь, овёс, просо, кукурузу, гречиху. В его кладовой можно обнаружить отдельные виды бобовых: горох, фасоль, сою. Запасы достигают внушительных размеров. Были найдены кладовые, в которых хранилось до 90 кг съестной продукции.
Весной поедает свежую зелень, корневища, корнеплоды. Летом делает набеги на сады, огороды, дачи. Примечательно, что хомячок никогда не съедает плод целиком, надгрызает один, а затем переходит к следующему. Таким способом он может испортить всю грядку лука, свёклы или моркови.
Летом делает набеги на сады, огороды, дачи. Примечательно, что хомячок никогда не съедает плод целиком, надгрызает один, а затем переходит к следующему. Таким способом он может испортить всю грядку лука, свёклы или моркови.
Помимо растительной пищи, питается падалью, рачками, насекомыми, моллюсками, мышами.
Размножение
Самки данного вида очень плодовиты, поэтому численность хомяков увеличивается из года в год. Половой зрелости достигают уже в 20 недель. В степных районах плодятся до четырёх раз, в горных – два раза. В одном помёте может рождаться до 18 детёнышей. Беременность длится от 15 до 22 дней. Малыши рождаются лысыми, слепыми, глухими. На седьмой день жизни их тельца покрываются шерстью, а через две недели они могут самостоятельно питаться. По истечении двадцати дней молодняк уходит от матери, чтобы начать самостоятельную жизнь. Они активно расселяются и роют норы. У глупых молодых хомячков много врагов. На них охотятся цапли, крысы, хорьки, барсуки.
Вред для сельскохозяйственных угодий
Большой вред хомяк наносит сельскому хозяйству, дачникам и огородникам. Постоянное размножение и умение приспосабливаться к любым погодным условиям способствуют быстрому освоению новых территорий и угодий. За ночь несколько хомяков могут полностью уничтожить посевы одного огорода. Поэтому перед людьми встал острый вопрос, как бороться с таким вредителем. Разработано несколько способов:
Постоянное размножение и умение приспосабливаться к любым погодным условиям способствуют быстрому освоению новых территорий и угодий. За ночь несколько хомяков могут полностью уничтожить посевы одного огорода. Поэтому перед людьми встал острый вопрос, как бороться с таким вредителем. Разработано несколько способов:
- Борьба с помощью животных. Способ не особо эффективный, ведь не каждая кошка способна справиться с такой крупной особью. Ей под силу только подрастающее поколение. Собака может противостоять хомяку, но при этом истопчет весь огород. К тому же существует опасность заражения домашнего животного инфекцией, которая может попасть в кровь при укусе грызуна.
- Затапливание нор. Сложность заключается в том, чтобы найти все входы в нору, ведь осторожный грызун очень хорошо их маскирует. В ином случае, этот способ не эффективен.
- Раскапывание нор. Способ опасный для человека и трудозатратный. Им практически не пользуются.
- Капканы. Основное в этом способе – знать, куда нужно поставить капкан.

- Отравленные приманки. Не факт, что приманку съест именно хомяк. Отравиться может любое животное, домашнее, в том числе.
- Ультразвуковые отпугиватели. Стоят дорого и не решают проблем с численностью вредителей. Не эффективны на большой территории.

Хомяк Радде внешне похож на своих сородичей – домашних ручных хомячков. Его вид вызывает умиление и восторг, но постоянное увеличение численности популяции вынуждает бороться с этим видом грызуна.
Национальный парк ”Эльбрус” описание и фото — Россия — Кавказ: Приэльбрусье
Статьи » Путешествия » Национальный парк ”Эльбрус” описание и фото — Россия — Кавказ: Приэльбрусье
 5em black}</style><a href=’https://www.youtube.com/embed/TF6TCg6xS1U?autoplay=1&fs=0&loop=1&modestbranding=1′><img src=’https://img.youtube.com/vi/TF6TCg6xS1U/hqdefault.jpg’ alt=’National Park ”Elbrus” description and photos — Russia — Caucasus: Prielbrusye’><button aria-label=’Watch’><svg version=’1.1′ viewBox=’0 0 68 48′><path d=’M66.52,7.74c-0.78-2.93-2.49-5.41-5.42-6.19C55.79,.13,34,0,34,0S12.21,.13,6.9,1.55 C3.97,2.33,2.27,4.81,1.48,7.74C0.06,13.05,0,24,0,24s0.06,10.95,1.48,16.26c0.78,2.93,2.49,5.41,5.42,6.19 C12.21,47.87,34,48,34,48s21.79-0.13,27.1-1.55c2.93-0.78,4.64-3.26,5.42-6.19C67.94,34.95,68,24,68,24S67.94,13.05,66.52,7.74z’ fill=’#f00′></path><path d=’M 45,24 27,14 27,34′ fill=’#fff’></path></svg></button></a>» allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»» title=»National Park ”Elbrus” description and photos — Russia — Caucasus: Prielbrusye»> 5em black}</style><a href=’https://www.youtube.com/embed/TF6TCg6xS1U?autoplay=1&fs=0&loop=1&modestbranding=1′><img src=’https://img.youtube.com/vi/TF6TCg6xS1U/hqdefault.jpg’ alt=’National Park ”Elbrus” description and photos — Russia — Caucasus: Prielbrusye’><button aria-label=’Watch’><svg version=’1.1′ viewBox=’0 0 68 48′><path d=’M66.52,7.74c-0.78-2.93-2.49-5.41-5.42-6.19C55.79,.13,34,0,34,0S12.21,.13,6.9,1.55 C3.97,2.33,2.27,4.81,1.48,7.74C0.06,13.05,0,24,0,24s0.06,10.95,1.48,16.26c0.78,2.93,2.49,5.41,5.42,6.19 C12.21,47.87,34,48,34,48s21.79-0.13,27.1-1.55c2.93-0.78,4.64-3.26,5.42-6.19C67.94,34.95,68,24,68,24S67.94,13.05,66.52,7.74z’ fill=’#f00′></path><path d=’M 45,24 27,14 27,34′ fill=’#fff’></path></svg></button></a>» allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»» title=»National Park ”Elbrus” description and photos — Russia — Caucasus: Prielbrusye»> Эльбрусский национальный парк описание и фото — Россия — Кавказ: Приэльбрусье. Фото и описаниеНациональный парк Эльбрус расположен на территории Кабардино-Балкарской Республики. Он был основан в 1986 с двумя основными задачами: создание оптимальных условий для развития туризма, рекреации, альпинизма и, конечно же, сохранение уникального природного комплекса. Парк расположен в пределах Зольского и Тырнаузского административных районов Кабардино-Балкарии. В границах парка «Приэльбрусье» расположены шесть населенных пунктов, в которых проживает более 6 тысяч человек. На территории парка произрастает около 400 видов растений. Особо охраняемым видом является рододендрон кавказский. В Красную книгу Российской Федерации занесены следующие растения: колокольчик доломитовый, береза Радде, граб хмельник обыкновенный, нут малый, камнеломка Динника. Береза Радде в 1885 г. была описана известным кавказским естествоиспытателем Г. Раде. Этот эндемичный реликтовый вид встречается лишь в некоторых районах Кавказа. Животный мир Эльбрусского парка тоже достаточно богат. Здесь обитает 111 видов птиц, более 60 видов млекопитающих, 8 видов амфибий, а также 11 видов рептилий, 6 видов рыб и множество видов насекомых. В парке обитают животные европейской степной зоны — хомяк обыкновенный, слепыш, серая куропатка, степной хорек и другие, и европейских лиственных лесов, среди них — косуля, европейский лесной кот, лесная куница и бурый медведь. Среди эндемиков Кавказа — кавказский тур, улар, тетерев, выдра и многие другие. Эльбрусский национальный парк — горнолыжный центр, экскурсионный центр отдыхающих на местных курортах, центр горно-спортивного туризма, место отдыха жителей и гостей Кабардино-Балкарской Республики. В парке 23 места отдыха. Рекомендуем также ознакомиться с описанием и фото Городского музея Осло (Oslo Bymuseum) — Норвегия: Осло Тема: Национальный парк ”Эльбрус” описание и фото — Россия — Кавказ: Приэльбрусье . Автор: Келли Костин |
 Подробная информация о аттракционе. Описание, фотографии и карта с указанием ближайших значимых объектов.
Подробная информация о аттракционе. Описание, фотографии и карта с указанием ближайших значимых объектов. Леса занимают лишь десятую часть всей территории Эльбрусского парка. Из лиственных пород наиболее распространены березы Радде и Литвинова (52,6%), из хвойных — сосна Коха (46,7%).
Леса занимают лишь десятую часть всей территории Эльбрусского парка. Из лиственных пород наиболее распространены березы Радде и Литвинова (52,6%), из хвойных — сосна Коха (46,7%).
Темы по странам
BK-каналы волосковых клеток взаимодействуют с RACK1, а PKC увеличивает свою экспрессию на клеточной поверхности посредством непрямого фосфорилирования
1.
Адамс Д.Р., Рон Д., Кили П.А.
RACK1, многогранный каркасный белок: структура и функция. Сигнал сотовой связи
9: 22, 2011 [бесплатная статья PMC] [PubMed] [Google Scholar]
2.
Алиуа А., Махаджан А., Нишимару К., Зарей М.М., Стефани Э., Торо Л.
Связь c-Src с большим напряжением проводимости и Ca 2+ -активированные K + каналы как новый механизм агонист-индуцированной вазоконстрикции. Proc Natl Acad Sci USA
99: 14560–14565, 2002 [бесплатная статья PMC] [PubMed] [Google Scholar]
3.
Амос С., Мартин П.М., Полар Г.А., Парсонс С.Дж., Хуссаини И.М.
Форбол-12-миристат-13-ацетат индуцирует трансактивацию рецептора эпидермального фактора роста посредством путей протеинкиназы Cdelta/c-Src в клетках глиобластомы. J Биол Хим
J Биол Хим
280: 7729–7738, 2005 [бесплатная статья PMC] [PubMed] [Google Scholar]
4.
Арт Джей Джей, Кроуфорд AC, Феттиплейс Р.
Электрический резонанс и мембранные токи в волосковых клетках улитки черепахи. Исследования слуха
22: 31–36, 1986 [PubMed] [Google Scholar]
5.
Арт Джей Джей, Феттиплейс Р.
Изменение свойств мембран волосковых клеток, выделенных из улитки черепахи. Журнал физиологии
385: 207–242, 1987 [бесплатная статья PMC] [PubMed] [Google Scholar]
6.
Art JJ, Fettiplace R, Wu YC.
Влияние низкого содержания кальция на зависящую от напряжения проводимость, связанную с настройкой волосковых клеток черепахи. Журнал физиологии
470: 109–126, 1993 [бесплатная статья PMC] [PubMed] [Google Scholar]
7.
Арт Дж.Дж., Ву Ю.С., Феттиплейс Р.
Активируемые кальцием калиевые каналы волосковых клеток черепахи. Журнал общей физиологии
105: 49–72, 1995 [бесплатная статья PMC] [PubMed] [Google Scholar]
8.
Бай Дж.П., Сургучев А., Джоши П., Гросс Л., Наваратнам Д.
CDK5 взаимодействует с Slo и влияет на его поверхностную экспрессию и кинетику посредством прямого фосфорилирования. Am J Physiol Cell Physiol
302: C766–C780, 2012 [бесплатная статья PMC] [PubMed] [Google Scholar]
9.
Бай Дж.П., Сургучев А., Наваратнам Д.
β 4 -Субъединица повышает чувствительность Slo к физиологическим концентрациям Ca 2+ и вместе с β 1 снижает поверхностную экспрессию Slo в волосковых клетках. Am J Physiol Cell Physiol
300: C435–C446, 2011 [бесплатная статья PMC] [PubMed] [Google Scholar]
10.
Берг Т., Бломхофф Р., Нэсс Л., Толлешауг Х., Древон К.А.
Монензин ингибирует рецептор-опосредованный эндоцитоз асиалогликопротеинов в гепатоцитах крыс. Разрешение ячейки опыта
148: 319–330, 1983 [PubMed] [Google Scholar]
11.
Бьян С., Бай Дж. П., Чапин Х., Ле Мёллик С., Донг Х., Каплан М., Сигворт Ф.Дж., Наваратнам Д.С.
Взаимодействия между бета-катенином и калиевым каналом HSlo регулируют поверхностную экспрессию HSlo. PLoS один
PLoS один
6: e28264, 2011 [бесплатная статья PMC] [PubMed] [Google Scholar]
12.
Блот В., Макгроу Т.Э.
Использование количественной иммунофлуоресцентной микроскопии для изучения внутриклеточного транспорта: исследования переносчика глюкозы GLUT4. Методы Мол Биол
457: 347–366, 2008 [PubMed] [Google Scholar]
13.
Ча С.К., Ву Т., Хуан С.Л.
Протеинкиназа С ингибирует опосредованный кавеолами эндоцитоз TRPV5. Am J Physiol Физиол почек
294: F1212–F1221, 2008 [PubMed] [Google Scholar]
14.
Чанг Б.И., Конрой К.Б., Махледер Э.М., Картрайт К.А.
RACK1, рецептор активированной С-киназы и гомолог бета-субъединицы G-белков, ингибирует активность тирозинкиназ src и рост клеток NIH 3T3. Мол Селл Биол
18: 3245–3256, 1998 [бесплатная статья PMC] [PubMed] [Google Scholar]
15.
Кроуфорд AC, Феттиплейс Р.
Механизм электрической настройки в волосковых клетках улитки черепахи. J Physiol (Лондон)
312: 377–412, 1981 [Бесплатная статья PMC] [PubMed] [Google Scholar]
16.
Диес-Сампедро А. , Сильверман В.Р., Баутиста Дж.Ф., Ричерсон Г.Б.
, Сильверман В.Р., Баутиста Дж.Ф., Ричерсон Г.Б.
Механизм повышенной вероятности открытия при мутации канала BK. J Нейрофизиол
96: 1507–1516, 2006 [PubMed] [Google Scholar]
17.
Du W, Bautista JF, Yang H, Diez-Sampedro A, You SA, Wang L, Kotagal P, Luders HO, Shi J, Cui J, Richerson GB, Wang QK.
Чувствительная к кальцию калиевая каналопатия при эпилепсии человека и пароксизмальном двигательном расстройстве. Нат Жене
37: 733–738, 2005 [PubMed] [Google Scholar]
18.
Дункан Р.К., Фукс П.А.
Различия в активируемых кальцием калиевых каналах с большой проводимостью от волосковых клеток вдоль базилярного сосочка курицы. Журнал физиологии
547: 357–371, 2003 [бесплатная статья PMC] [PubMed] [Google Scholar]
19.
Фернандес-Фернандес Дж.М., Томас М., Васкес Э., Орио П., Латорре Р., Сенти М., Марругат Дж., Вальверде М.А.
Мутация усиления функции в субъединице калиевого канала KCNMB1 связана с низкой распространенностью диастолической гипертензии. Джей Клин Инвест
113: 1032–1039, 2004 [бесплатная статья PMC] [PubMed] [Google Scholar]
20.
Феттиплейс Р., Фукс П.А.
Механизмы настройки волосковых клеток. Ежегодный обзор физиологии
61: 809–834, 1999 [PubMed] [Google Scholar]
21.
Frucht CS, Uduman M, Kleinstein SH, Santos-Sacchi J, Navaratnam DS.
Градиенты экспрессии генов вдоль тонотопической оси слухового эпителия кур. J Assoc Res Отоларингол
12: 423–435, 2011 [бесплатная статья PMC] [PubMed] [Google Scholar]
22.
Гриффитс Г., Куинн П., Уоррен Г.
Рассечение комплекса Гольджи. I. Монензин ингибирует транспорт белков вирусной мембраны от медиальной к транс-цистерне Гольджи в клетках почек детенышей хомячка, инфицированных вирусом леса Семлики. Джей Селл Биол
96: 835–850, 1983 [бесплатная статья PMC] [PubMed] [Google Scholar]
23.
Хадспет А.Дж., Льюис Р.С.
Модель электрического резонанса и настройки частоты в мешотчатых волосковых клетках лягушки-быка Rana catesbeiana. Журнал физиологии
400: 275–297, 1988 [бесплатная статья PMC] [PubMed] [Google Scholar]
24.
Исаксон К.К., Лу Кью, Карас Р.Х., Кокс Д. Х.
Х.
RACK1 представляет собой белок, связывающий канал BK Ca . Am J Physiol Cell Physiol
292: C1459–C1466, 2007 [PubMed] [Google Scholar]
25.
Исса Н.П., Хадспет А.Дж.
Кластеризация Ca 2+ каналов и Са 2+ -активированных каналов К + на флуоресцентно меченных пресинаптических активных зонах волосковых клеток. Proc Natl Acad Sci USA
91: 7578–7582, 1994 [бесплатная статья PMC] [PubMed] [Google Scholar]
26.
Исса Н.П., Хадспет А.Дж.
Поступление и клиренс Ca 2+ в отдельных пресинаптических активных зонах волосковых клеток из саккулюса лягушки-быка. Proc Natl Acad Sci USA
93: 9527–9532, 1996 [бесплатная статья PMC] [PubMed] [Google Scholar]
27.
Карпова Т.С., Бауманн К.Т., Хе Л., Ву С., Грэммер А., Липски П., Хагер Г.Л., МакНалли Дж.Г.
Резонансный перенос энергии флуоресценции от голубого к желтому флуоресцентному белку, обнаруженный фотообесцвечиванием акцептора с использованием конфокальной микроскопии и одного лазера. Дж Микроск
Дж Микроск
209: 56–70, 2003 [PubMed] [Google Scholar]
28.
Ким Дж.М., Бейер Р., Моралес М., Чен С., Лю Л.К., Дункан Р.К.
Экспрессия вариантов сплайсинга калиевых каналов, активируемых кальцием BK-типа, во время развития улитки цыпленка. Дж Комп Нейрол
518: 2554–2569, 2010 [бесплатная статья PMC] [PubMed] [Google Scholar]
29.
Кок К., Коенен А., Гизе Б., Фраунхольц М., Мэй К., Зигмунд В., Хаммер Э., Фолькер У., Едлички Г., Кремер Х.К., Грубе М.
Быстрая модуляция функции полипептида 2B1, транспортирующего органические анионы (OATP2B1, SLCO2B1), посредством интернализации, опосредованной протеинкиназой C. J Биол Хим
285: 11336–11347, 2010 [бесплатная статья PMC] [PubMed] [Google Scholar]
30.
Ли С., Эстерберг Р., Лашанс В., Рен Д., Радде-Галлвитц К., Чи Ф., Парент Дж.Л., Фриц А., Чен П.
Rack1 необходим для локализации мембраны Vangl2 и передачи сигналов планарной клеточной полярности при ослаблении канонической активности Wnt. Proc Natl Acad Sci USA
108: 2264–2269, 2010 [бесплатная статья PMC] [PubMed] [Google Scholar]
31.
Манна П.Т., Смит А.Дж., Танеджа Т.К., Хауэлл Г.Дж., Липпиат Д.Д., Шивапрасадарао А.
Конститутивная эндоцитарная рециркуляция и опосредованная протеинкиназой С лизосомальная деградация контролируют поверхностную плотность канала K ATP . J Биол Хим
285: 5963–5973, 2010 [бесплатная статья PMC] [PubMed] [Google Scholar]
32.
Миранда-Роттманн С., Козлов А.С., Хадспет А.Дж.
Высокоспецифичный альтернативный сплайсинг транскриптов, кодирующих каналы BK в улитке кур, является минорной детерминантой тонотопического градиента. Мол Селл Биол
30: 3646–3660, 2010 [бесплатная статья PMC] [PubMed] [Google Scholar]
33.
Наваратнам Д., Бай Дж. П., Самаранаяке Х., Сантос-Сакки Дж.
Опосредованная N-концом гомомультимеризация престина, моторного белка наружных волосковых клеток. Биофиз Дж
89: 3345–3352, 2005 [бесплатная статья PMC] [PubMed] [Google Scholar]
34.
Наваратнам ДС.
Двухгибридный скрининг дрожжей для проверки белок-белковых взаимодействий в слуховой системе. Методы Мол Биол
493: 257–268, 2009 [PubMed] [Google Scholar]
35.
Нельсон МТ, Бонев АД.
Субъединица бета1 Ca 2+ -чувствительного K 9Канал 0043+ защищает от гипертонии. Джей Клин Инвест
113: 955–957, 2004 [бесплатная статья PMC] [PubMed] [Google Scholar]
36.
Плугер С., Фаулхабер Дж., Фюрстенау М., Лон М., Вальдшютц Р., Голлаш М., Халлер Х., Люфт Ф.К., Эмке Х., Понгс О.
Мыши с нарушенным геном субъединицы бета1 канала BK имеют аномальную связь Ca 2+ искра/STOC и повышенное кровяное давление. Цирк Рез
87: E53–E60, 2000 [PubMed] [Google Scholar]
37.
Пиотт С.Дж., Мередит А.Л., Фодор А.А., Васкес А.Е., Ямоа Э.Н., Олдрич Р.В.
Кохлеарная функция у мышей, лишенных субъединиц альфа, бета1 или бета4 канала BK. J Биол Хим
282: 3312–3324, 2007 [PubMed] [Google Scholar]
38.
Рон Д., Чен Ч., Колдуэлл Дж., Джеймисон Л., Орр Э., Мочли-Розен Д.
Клонирование внутриклеточного рецептора протеинкиназы С: гомолога бета-субъединицы G-белков. Proc Natl Acad Sci USA
91: 839–843, 1994 [бесплатная статья PMC] [PubMed] [Google Scholar]
39.
Sailer CA, Kaufmann WA, Kogler M, Chen L, Sausbier U, Ottersen OP, Ruth P, Shipston MJ, Knaus HG.
Иммунолокализация каналов BK в пирамидных нейронах гиппокампа. Eur J Neurosci
24: 442–454, 2006 [PubMed] [Google Scholar]
40.
Самаранаяке Х., Сондерс Дж. К., Грин М. И., Наваратнам Д. С.
Ca 2+ и K + (BK) каналы в волосковых клетках кур сгруппированы и колокализованы с апикально-базальным и тонотопическим градиентами. Дж Физиол
560: 13–20, 2004 [бесплатная статья PMC] [PubMed] [Google Scholar]
41.
Serrels B, Sandilands E, Serrels A, Baillie G, Houslay MD, Brunton VG, Canel M, Machesky LM, Anderson KI, Frame MC.
Комплекс между FAK, RACK1 и PDE4D5 контролирует инициацию распространения и полярность раковых клеток. Карр Биол
20: 1086–1092, 2010 [PubMed] [Google Scholar]
42.
Шипстон М.Дж., Армстронг Д.Л.
Активация протеинкиназы С ингибирует активируемые кальцием калиевые каналы в опухолевых клетках гипофиза крыс. Дж Физиол
493: 665–672, 1996 [бесплатная статья PMC] [PubMed] [Google Scholar]
43.
Соколовский Б., Орчард С., Харви М., Шридхар С., Сакаи Ю.
Исправление: взаимодействия консервативных каналов BK с белками выявляют сигналы, относящиеся к гибели и выживанию клеток. PLoS один
7: 2011 [бесплатная статья PMC] [PubMed] [Google Scholar]
44.
Торнело К.С., Нельсон М.Т.
Ионные каналы в гладких мышцах: регуляторы внутриклеточного кальция и сократимости. Can J Physiol Pharmacol
83: 215–242, 2005 [PubMed] [Google Scholar]
45.
Тиан Л., Когхилл Л.С., Макклафферти Х., Макдональд С.Х., Антони Ф.А., Рут П., Кнаус Х.Г., Шипстон М.Дж.
Различная стехиометрия фосфорилирования тетрамера канала BK Ca указывает на активацию и ингибирование канала цАМФ-зависимой протеинкиназой. Proc Natl Acad Sci USA
101: 11897–11902, 2004 [бесплатная статья PMC] [PubMed] [Google Scholar]
46.
Учида Н., Смиловиц Х., Танцер М.Л.
Моновалентные ионофоры ингибируют секрецию проколлагена и фибронектина из культивируемых фибробластов человека. Proc Natl Acad Sci USA
76: 1868–1872, 1979 [бесплатная статья PMC] [PubMed] [Google Scholar]
47.