Различие полов у рыб. Есть ли у рыб пол
Различие полов у рыб
По биологическому значению функция размножения у животных уступает только добыванию корма. Однако в период высокой половой активности половая доминанта подавляет и чувство голода. В водной среде процесс размножения отличается относительной простотой по технике исполнения. Чаще всего оплодотворение в воде носит вероятностный характер, а потому количество вариантов должно быть достаточно большим, чтобы сделать процесс оплодотворения результативным. Поэтому в целом для класса рыб характерна высокая плодовитость. Количество женских гамет за один период икрометания у представителей пелагических видов рыб достигает десятков и даже сотен миллионов, а количество сперматозоидов и того больше.
Среди животных с таким внутриклассовым морфофункциональным разнообразием, как рыбы, всегда есть исключения из общего правила. Это справедливо и по отношению к процессу размножения.
Существует ряд общих требований, без которых размножение невозможно. Во-первых, рыбы должны достичь возраста полового созревания. Правда, у рыб понятие "возраст полового созревания" достаточно расплывчато и не так привязано к календарному возрасту, как у высших позвоночных.
Межвидовые различия сроков полового созревания у рыб огромны: от 1 мес у гамбузии до 30 лет у некоторых видов осетровых рыб. У рыб сроки полового созревания помимо календарного возраста определяет ряд факторов внешней среды (температура воды, химический состав воды, интенсивность питания, плотность посадки в искусственные водоемы). Время полового созревания коррелирует с живой массой рыб.
Сроки полового созревания различаются у рыб разных популяций одного вида. Например, у леща половая зрелость наступает в популяции Ладожского озера в 8-9 лет, в популяции леща Средней Волги -в 6-7 лет, в Северном Каспии - в 4-6 лет. Азово-черноморская популяция леща начинает нереститься уже в 3 года. То же можно сказать и о других видах рыб. Например, у волжского и азовского осетров половая зрелость наступает в 9-10 лет. У куринского осетра половая зрелость самок наступает лишь в 30 летнем возрасте.
Половая дифференциация у рыб - явление уникальное. Рыбы обладают очень пластичной системой репродукции, которая позволила им занять практически все существующие на Земле водоемы. В соответствии с биологической необходимостью рыбы могут изменять половую принадлежность отдельных особей. У рыб нет единообразной хромосомной системы полового детерминизма, как, например, у человека (XX-, XY-хромосомы. т. е. системы половых гетерохромосом). Точнее, как вариант она встречается в классе рыб, например у рыбца. Самки рыбца в диплоидном наборе имеют две одинаковые половые хромосомы (XX), самцы рыбца имеют две разные половые хромосомы (XY).
У других видов, например японского угря, картина прямо противоположная: у самца имеется пара одинаковых половых хромосом (ZZ), а у самки -две разные половые хромосомы (WZ). Кроме того, есть виды, у которых у одного из полов половая хромосома непарная. Так, например, самка фундулюса имеет парные половые хромосомы (XX), а самец - непарную половую хромосому (ХО).
У лабиринтовой рыбы коллизии только самец несет половую хромосому, да и ту непарную.
У основных объектов рыборазведения - карповых, лососевых, представителей осетровых рыб -половых хромосом нет. У этих рыб половой детерминизм имеет полихромосомную основу, т. е. гены, кодирующие первичные и вторичные половые признаки, рассредоточены по другим соматическим хромосомам. Такие резкие различия полового генно-хромосомного детерминизма у рыб едва ли связаны с филогенезом, так как они встречаются в пределах одного семейства и даже в пределах одного вида (последнее характерно для разных популяций угрей и пециллий).
Неопределенность с половой принадлежностью остается и в онтогенезе. Чаще всего у молоди нет половых различий до момента полового созревания. При этом даже гистологические исследования гонад не проясняют ситуации с полом. У рыб первичные половые клетки способны развиваться как по Женскому, так и по мужскому пути. Многим видам рыб свойствен ювенальный гермафродитизм, т. е. параллельное развитие и мужских, и женских гонад и соответственно половых клеток, одни из которых впоследствии отмирают.
У рыб известно явление проандрии и прототинии, когда на раннем этапе онтогенеза рыбы развиваются мужские гонады, а на Последующем - женские. Как нормальное физиологическое явление у рыб встречается несколько типов функционального гермафродитизма. Особенно много примеров этого явления у окуневых рыб. Есть виды окуней, которые первую половину репродуктивного периода являются самками, а вторую- самцами. При этом рыбы имеют и первичные, и вторичные половые признаки с нормальным оно- или сперматогенезом и соответствующим полу нерестовым поведением.
И уж совсем необычно выглядит истинный суточный транссексуализм у морских окуней. В течение суток одна и та же особь многократно меняет свою половую принадлежность, выметывает или икру, или молоки. Самооплодотворение здесь маловероятно, но возможно.
Вторичные половые признаки рыб, которые особенно ярко проявляются в период нереста, имеют важное практическое значение в рыбоводстве при сортировке рыб по половому признаку.
Например, у лососей признаком самцов являются более яркая окраска и изменения опорно-двигательного аппарата - искривление челюстей, появление горба. У самца колюшки перед нерестом брюшко окрашивается в алый цвет. Самцы многих морских бычков перед нерестом становятся абсолютно черными. У карповых (вобла, язь) на голове и теле самцов в преднерестовый период появляется "жемчужная сыпь" - роговые образования белесого цвета. Появление брачного наряда у рыб определяется гормональными перестройками в этот период жизни. Значение его специалисты трактуют по-разному. Брачная окраска в нерестовый период может выполнять защитную роль. У лососевых брачный наряд маскирует рыб, делает их менее заметными на галечном грунте в прозрачной воде. В других случаях нерестовая окраска имеет сигнальное значение (горбуша, кета).
Пол одних рыб можно определить по форме анального отверстия, других - по форме плавников (рис. 11.1).
|
|
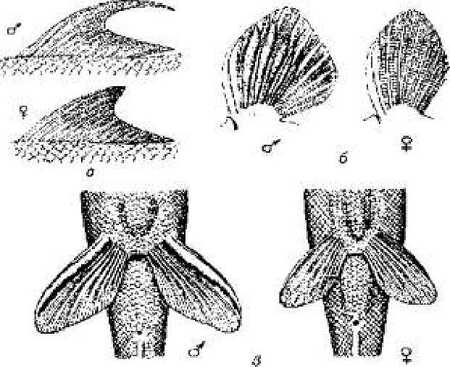
Рис. 11.1. Вторичные половые признаки рыб:
а-спинные плавники Lafaeo dero; б-грудные плавники тибетского гольца; в - брюшные плавники линя
Например, у живородящих пициллид половой диморфизм довольно рано проявляется в особом строении анального плавника самцов, выполняющего роль совокупительного органа. У акул такую же роль выполняет придаток брюшного плавника.
Важным половым признаком может служить половое поведение рыб перед нерестом. Это может быть преследование самцом самки (карп, щука и др.) или более сложное поведение - устройство гнезда (лабиринтовые), охрана территории. После нереста у многих видов рыб наблюдаются элементы родительского поведения: вентиляция икры, инкубация икры в ротовой полости, охрана молоди (тиляпия). Конечно, какими бы сложными формами поведения рыбы не обладали в период размножения, все они являются результатом врожденных стереотипических поведенческих актов. Половое поведение проявляется при изменении гормонального статуса рыб в ответ на действие внешней среды. Половая доминанта формируется под влиянием усиления функции гипоталамо-гипофизарно-гонадо-адреналовой системы.
Управление половым детерминизмом при искусственном разведении рыб имеет большое практическое значение. Особенно полезным этот прием может быть при разведении ценных рыб - осетровых, лососевых. Здесь желательно иметь большое поголовье самок и ограниченное количество самцов, однако половые признаки у этих видов проявляются довольно поздно, когда уже затрачено много средств и времени.
Регулировать пол в искусственных условиях выращивания удается при помощи стероидных гормонов. Андрогены и эстрогены не разрушаются в желудочно-кишечном тракте. Поэтому они добавляются в корма. Включения метилтестостерона в рацион личинок тиляпии в количестве 30-50 мг/кг приводит к тому, что в стаде половозрелых рыб самцы составляют 95-100 %. При добавлении в рацион форели этого же гормона (3 мг/кг) все особи превращались в самцов. При добавлении в рацион гормона эстрадиола (20 мг/кг корма) у лососей формировалось полностью (на 100 %) женское гомосексуальное стадо. Такой же "феминистический" эффект получали у тиляпии при помощи этинилэстрадиола в количестве 50 мг/кг корма.
Таким образом, гормональная регуляция пола у рыб при помощи синтетических или природных стероидов может претендовать на самостоятельный технологический прием в рыбоводстве.
biofile.ru
Определение пола у рыб | Cell Biology.ru
Определение пола.
У всех позвоночных животных пол индивидуума определяется генетически, то есть комбинацией половых хромосом, доставшихся от каждого из родителей при оплодотворении яйцеклетки. Но кроме генетических факторов, на определение пола у рыб могут влиять также температура и соленость воды, в которой развивается зародыш, соотношение периодов света и темноты. Определение пола под влиянием внешних условий называется фенотипическим, или модификационным.
Atlantic silversideТак David O. Conover (David O. Conover, 1984) пишет, что большинство рождающихся ранней весной, при пониженных температурах, атерин (Atlantic silverside) становятся самками. А рыбы, развивающиеся позже при более высокой температуре, вырастают самцами. Это имеет огромное значение для выживаемости данного вида. Численность потомства напрямую зависит от плодовитости самок. Так как в данном случае самки растут дольше самцов, они имеют большой размер и соответственно могут дать много икры, поскольку размер их половых органов пропорционаленразмеру тела, чего не наблюдается у самцов.
Варианты генетической детерминации пола.
Самцы с одной Х-хромосомой или с двумя разными (XY) хромосомами имеют гетерогаметный пол, самки с ХХ-хромосомами — гомогаметный пол. У многих животных, наоборот, самки имеют гетерогаметный пол. Их половые хромосомы обозначают буквами Z и W или XY, а половые хромосомы гомогаметных самцов - ZZ или XX. У млекопитающих, нематод, моллюсков, иглокожих и у большинства членистоногих гетерогаметен мужской пол. У насекомых и рыб гетерогаметность наблюдается как у мужского, так и у женского пола. Гетерогаметность женского пола свойственна птицам, пресмыкающимся и некоторым земноводным. При определении пола у рыб оказалось, что гуппи, пецилия, сфенопс, медака и др. принадлежат к типу ХХ- XY (мужская гетерогаметность). A platyfish Xiphophorus maculatus имеют как мужскую (XY), так и женскую гетерогаметность (WY) и даже три типа половых хромосом (WY, WX, XX).Разные линии мозамбикских тиляпий имеют гетерогаметностьсамок и самцов. Скрещивание этих линий привело к образованию в потомстве одних самцов. У зеленых меченосцев (X. helleri) и черных макроподов (М. ореrcularis concolor) дифференцировка пола зависит от мужских и женских наследственных факторов, расположенных в аутосомах (половых хромосом нет).Механизм генетического контроля над развитием половых признаков может быть внутри- и межклеточным. Внутриклеточное определение пола не связано с образованием половых гормонов (напр., у насекомых), и действие генов, определяющих пол, ограничено клетками, в которых эти гены функционируют. При этом в одном организме могут нормально развиваться, не влияя друг на друга, участки тела с женскими и мужскими признаками. При межклеточном определении пола, характерном для млекопитающих и птиц, под контролем генов вырабатываются половые гормоны, которые, проникая во все клетки организма, обусловливают фенотипическое развитие признаков соответствующего пола. Различают прогамное, сингамное и эпигамное определение пола.Прогамное определение пола происходит до оплодотворения яйца, напр. дифференцировка яйцеклеток на быстро и медленно растущие. Первые становятся крупными, и из них после оплодотворения развиваются самки, вторые отличаются меньшими размерами и дают самцов, хотя оба вида яйцеклеток генетически одинаковы. Сингамное определение пола происходит во время оплодотворения, но на разных стадиях этого процесса. У некоторых видов с мужской гетерогамией и физиологической полиспермией (оплодотворение яйцеклетки несколькими сперматозоидами) пол определяется в момент кариогамии. Если с ядром яйцеклетки сливается мужское ядро с Y-хромосомой, разовьётся мужская особь, если с Х-хромосомой – женская. При женской гетерогамии пол потомства зависит от того, какая из половых хромосом попадает в ядро яйцеклетки во время мейоза. Если в ядре окажется Z-хромосома, разовьётся особь мужского пола, если W-хромосома – женского. Т.о, в данном случае пол зиготы устанавливается до кариогамии.Эпигамное определение пола наблюдается у разнополых видов с фенотипическим определением пола, когда направленность развития в сторону мужского или женского пола обусловливается влиянием внешних условий после оплодотворения.
Эндокринная регуляция дифференцировки пола у рыб.
В процессе дифференцировки пола у разнополых рыб центральная нервная система (ЦНС) участвует непродолжительное время (Baroiller, 1999). Вместе с тем, в течение этого времени в работе задействована гипоталамо-гипофизарная система посредством гонадотропир-релизинг фактора (ГРФ) и гонадотропиновых (ГТ) гормонов. При использовании специфических антител в гипофизе rainbow trout были найдены ГТ1 иммуночувствительные клетки на стадии, когда митоз эмбриональных клеток был впервые зарегистрирован в половых железах (Saga et al, 1993). Иммуноспецифичность к ГРФ наблюдалась только в течение половой дифференцировки, причем у обоих полов и в сходных отделах мозга, включая те, где обнаруживалсяГТ1. Чувствительность к гормону роста (ГР) в гипофизе обнаруживалась на более ранних стадиях чем к ГТ1. Было высказано предположение, что ГР может быть вовлечен в процесс дифференцировки пола, так как известно, что он участвует в регуляции синтеза стероидов в половых железах взрослых рыб. Иначе говоря, гонадотропин стимулирует продукцию стероидов, но уже после гистологической дифференцировки гонад. На более ранних стадиях была отмечена стимуляция продукции андростиндиона межпочкой. Не известно точно связана ли эта продукция стероидов с дифференцировкой гонад напрямую, но ясно, что участие межпочки в синтезе стероидов, которые в свою очередь влияют на дифференцировку гонад, помогает становлению пола у рыб. У гермофродитных видов рыб роль ЦНС имеет большое значение (Baroiller, 1999). Гонадотропины играют большую роль в инициации процесса инверсии пола. Это может быть связано с социальным контролем инверсии пола, который демонстрируется многими видами гермафродитов. Ключевым ферментом является ароматаза,которая превращает андрогены в эстрогены и тем самым способствует дифференцировке или инверсии пола. Также у рыб были найдены некоторые гены млекопитающих, которые имеют большое значение в процессе дифференцировки пола у последних (Hofsten and Olsson, 2005). Но у рыб их роль пока не изучена достаточно хорошо. К числу таких генов относятся: Sox9a and Sox9b, AMH, WT1, Drosophila fushi tarazu factor-1 (FTZ-F1).К примеру, у zebrafish не известны механизмы, контролирующие детерминацию и дифференцировку пола. Также не идентифицировано ни одного гена, связанного с полом и ни одной половой хромосомы. Тем не менее, некоторое число генов обнаруживает связь с процессом дифференцировки и детерминации пола. В центре всеобщего внимания сейчас находятся FTZ-F1 гены zebrafish, так как они вовлечены в развитие межпочки, а, следовательно, и в биосинтез стероидов, так как обнаруживают экспрессию, совпадающую с дифференцировкой и функционированием репродуктивной ткани (Hofsten and Olsson, 2005). Zebrafish может менятьпол при воздействии на нее эстрогенами, что наводит на мысль о решающей рола уровня эстрогенов в процессе дифференцировки пола. Продукт гена Cyp19 является ароматазой, обращающей тестостерон в 17-?-эстрадиол, и самцов (при большом их числе) в самок.
Соотношение полов.
При фенотипическом определении пола оно зависит от количества развивающихся организмов, которые попадают под влияние внешних факторов, детерминирующих тот или иной пол. При генетическом определении пола соотношение полов у большинства видов, как правило, очень близко к соотношению: 100 самок на 100 самцов. Однако и при таком определении пола есть отклонения. Так, у некоторых видов млекопитающих с мужской гетерогамией статистически достоверно рождается на 1 -2% больше потомков мужского пола. Изменение соотношения полов может быть почти у всех животных с генетическим определением пола результатом гибели половины зародышей гетерогаметного пола под действием сцепленных с полом леталей или рецессивных мутаций, несовместимых с жизнью.
Реверсия пола.
Существенный сдвиг соотношения организмов в сторону одного из полов имеет как теоретическое, так и практическое значение, т. к. один из полов обычно более продуктивен. Методы регуляции пола применяются в зависимости от типа определения пола и биологических и хозяйственных особенностей вида. При фенотипическом переопределении пола, если действие генов реализуется посредством гормонов, половые признаки изменяются при пересадке половых органов одного пола другому или при введении в организм гормонов противоположного пола, а также некоторых аминокислот. Степень фенотипических изменений пола зависит от особенностей вида и дозы введённого препарата. Однако лишь в редких случаях (у некоторых рыб и земноводных) особи с фенотипически переопределённым полом продуцируют гаметы, противоположные их генотипическому полу. В следующем поколении, если действие гормонов прекращается, снова вступает в силу генетический механизм определения пола.Уже давно было замечено, что самки рыб под воздействиемопределенных условий (среды, гормональных препаратов и т. п.) превращаются в самцов (гамбузиевые, меланохромисы и т. д.). Женский половой гормон (эстрол) и мужской (метилтестостерон) при добавлении в воду или корм переопределяют пол. Японский генетик Ямамото (Yamamoto, 1969) превратил самок золотых рыбок в самцов. При дальнейшем разведении все потомство оказалось женским. На сегодняшний день рыбоводы научились менять пол рыб, воздействуя небольшими дозами гормонов на икру.Также повреждение эндокринной системы самих рыб может стать причиной преимущественного рождения особей одного пола. Так Ларссон (Larsson at all, 2000) пишет, что вредные выбросы промышленных заводов вызывают нарушения в эндокринной системе рыб на ранних стадиях развития, что резко меняет соотношение полов в сторону преимущественного развития самцов.Но самое интересное – это социальные факторы изменения пола – то есть взрослые половозрелые рыбы меняют пол под влиянием «общественного мнения» так, чтобы оптимизировать соотношениеполов в популяции. Что же дает рыбе смена пола? Установлено, что количество и качество икры прямо зависит от размера самки. Для самцов такой зависимости нет. Поэтому стратегия, при которой рыбы сначала вступают в размножение как самцы, а став старше и крупнее, превращаются в самок, выглядит логичной. Так обстоит дело у дениса, у которого большая часть молодых рыб нерестится сначала как мелкие самцы, а через год уже как крупные самки. Превращение самцов в самок под влиянием социальных факторов хорошо изучено на примере рыбы-клоуна. Молодь клоуна оседает из планктона на риф под защиту актиний. Самая крупная рыба в группе становится самкой, вторая по величине – самцом. Остальные рыбы остаются неполовозрелыми. Но если самка погибает, то клоун-самец быстро превращается в самку, а следующая по величине рыба созревает как самец и занимает его место. Однако бывают ситуации, когда успех в размножении зависит от размеров самца. В случаях, когда самцу необходимо защищать гарем, нерестовую территорию илиохранять кладку икры, крупные самцы имеют преимущество перед мелкими. Поэтому у некоторых видов самки растут-растут и превращаются в самцов. Например, рыба шури (император) сначала нерестится как самка, а в следующий раз – как самец.Но есть на коралловых рифах рыбы, которым не нужно менять пол, поскольку они одновременно являются самцами и самками. У таких рыб-гермафродитов созревают и икра, и молоки. Если две такие особи встретятся и понравятся друг другу, то каждая из них принесет потомство. Если же достойного партнера не найдется, то можно прибегнуть и к самооплодотворению. Такая стратегия дает преимущество малочисленным глубоководным рыбам, которым трудно найти себе пару в океанских глубинах. Но до сих пор совершено непонятно, зачем это нужно коралловым рыбам. Коралловые рыбы-гермафродиты живут достаточно плотными популяциями, в которых выбор пары – не проблема. Более того, известны виды гермафродитов, которые образуют постоянные моногамные пары на всю жизнь.
Эволюция пола.
Раздельнополость (бисексуальность), свойственная уже многим одноклеточным (водоросли, простейшие), произошла от смешаннополости. Лишь в некоторых случаях (напр., при паразитизме) смешаннополость могла возникнуть вторично из раздельнополости. Так, у паразитических ракообразных наблюдаются все переходы от смешанно- к раздельнополости (напр., раздельнополые виды с хорошо развитыми самками и карликовыми самцами – явственный сдвиг в сторону гермафродитизма). Фенотипическое определение пола древнее генетического, т. к. на ранних стадиях эволюции ещё не существовало специального аппарата половых хромосом. Возникающие на определённых этапах эволюции специальные половые хромосомы (рыбы, земноводные) первоначально морфологически неотличимы от аутосом, и о наличии их можно судить только по признакам, сцепленным с полом. Вслед за морфологическими различиями между половыми хромосомами и аутосомами возникает дифференцировка между Х- и Y-хромосомой, что делает всё более редкой конъюгациюмежду ними и затрудняет обмен их участками при кроссинговере. Всё это способствует выполнению специфических функций половых хромосом – быть реализаторами женского или мужского пола. Полное исчезновение Y-хромосомы делает генетическое определение пола ещё более совершенным: пол определяется равновесием между числом аутосом и хромосом.Так Уолтер Трот и Хенз Уинкин (Traut and Winking, 2001) сообщили о результатах исследования мейотических хромосом и стадий эволюции половых хромосом на примере таких рыб как zebrafish, platyfish и guppy. После сравнительной геномной гибридизации митотических и мейотических хромосом этих трех видов было обнаружено, что они демонстрируют основные ступени дифференцировки половых хромосом. А именно: zebrafish имеет кориотип, состоящий из одних аутосом, platyfish – с генетически детерминированными половыми хромосомами, которые еще не подразделяются на X и Y-хромосомы, и, наконец, guppy с генетически и цитогенетически дифференцированными половыми хромосомами.Для zebrafish характерна половая дифференцировка в зависимости от условий окружающей среды. Для этого вида половые хромосомы не идентифицируются с помощью C-banding, хромомицина А3 или репликативного бэндинга. На стадии пахитены и диакинеза ни один из 25 бивалентов не демонстрирует морфологическую и молекулярную дифференцировку половых хромосом. Platyfish Xiphophorus maculatus имеют четко определенную систему половых хромосом, но с некоторыми вариациями между различными популяциями рыб. Так мексиканская популяция имеет XX/XY систему определения пола, в то время как некоторые другие популяции имеют W-хромосомы помимо X и Y. Хромосомы X и Y не имеют видимых отличий. Молекулярный маркер XIR, который был локализован на дистальном конце одной из акроцентрических хромосом, идентифицировал ее как Y-хромосому (Nanda et al. 2000). В том районе также находится пол-определяющий район (SDR – sex determining region). У guppy акроцентрическая Y-хромосома состоит изпроксимального сегмента, гомологичного району на X-хромосоме, с которым они спариваются на стадии ранней пахитены, и дистального сегмента, отличного от сегментов Х-хромосомы, который неспаривается с Х-хромосомой на стадии ранней пахитены, но образует синапсы позже. Он включает в себя уже известный пол-определяющий район и бросающийся в глаза вариабельный гетерохроматиновый район, чья структура зависит от индивидуальной линии Y-хромосомы. Также сравнительная геномная гибридизация показала большой блок преимущественно самец-специфичной повторенной ДНК (Nanda et al. 1990) и блок общей неспецифической ДНК. Так после гибридизации с олигонуклеотидной пробой (GACA)4 был обнаружен самец-специфичный локус повторов, который отличал Y-хромосому самцов с красным цветом тела.Болтенгаген A.A.
www.cellbiol.ru
Дифференциация полов у рыб » Рыбалка на Урале
Половая дифференциация у рыб - явление уникальное. Рыбы обладают очень пластичной системой репродукции, которая позволила им занять практически все существующие на Земле водоемы. В соответствии с биологической необходимостью рыбы могут изменять половую принадлежность отдельных особей. У рыб нет единообразной хромосомной системы полового детерминизма, как, например, у человека (XX-, XY-хромосомы. т. е. системы половых гетерохромосом). Точнее, как вариант она встречается в классе рыб, например у рыбца. Самки рыбца в диплоидном наборе имеют две одинаковые половые хромосомы (XX), самцы рыбца имеют две разные половые хромосомы (XY).У других видов, например японского угря, картина прямо противоположная: у самца имеется пара одинаковых половых хромосом (ZZ), а у самки -две разные половые хромосомы (WZ). Кроме того, есть виды, у которых у одного из полов половая хромосома непарная. Так, например, самка фундулюса имеет парные половые хромосомы (XX), а самец - непарную половую хромосому (ХО).
У лабиринтовой рыбы коллизии только самец несет половую хромосому, да и ту непарную. У основных объектов рыборазведения -- карповых, лососевых, представителей осетровых рыб -половых хромосом нет. У этих рыб половой детерминизм имеет полихромосомную основу, т. е. гены, кодирующие первичные и вторичные половые признаки, рассредоточены по другим соматическим хромосомам. Такие резкие различия полового генно-хромосомного детерминизма у рыб едва ли связаны с филогенезом, так как они встречаются в пределах одного семейства и даже в пределах одного вида (последнее характерно для разных популяций угрей и пециллий).
Неопределенность с половой принадлежностью остается и в онтогенезе. Чаще всего у молоди нет половых различий до момента полового созревания. При этом даже гистологические исследования гонад не проясняют ситуации с полом. У рыб первичные половые клетки способны развиваться как по Женскому, так и по мужскому пути. Многим видам рыб свойствен ювенальный гермафродитизм, т. е. параллельное развитие и мужских, и женских гонад и соответственно половых клеток, одни из которых впоследствии отмирают.
У рыб известно явление проандрии и прототинии, когда на раннем этапе онтогенеза рыбы развиваются мужские гонады, а на Последующем - женские. Как нормальное физиологическое явление у рыб встречается несколько типов функционального гермафродитизма. Особенно много примеров этого явления у окуневых рыб. Есть виды окуней, которые первую половину репродуктивного периода являются самками, а вторую- самцами. При этом рыбы имеют и первичные, и вторичные половые признаки с нормальным оно- или сперматогенезом и соответствующим полу нерестовым поведением.
И уж совсем необычно выглядит истинный суточный транссексуализм у морских окуней. В течение суток одна и та же особь многократно меняет свою половую принадлежность, выметывает или икру, или молоки. Самооплодотворение здесь маловероятно, но возможно. Вторичные половые признаки рыб, которые особенно ярко проявляются в период нереста, имеют важное практическое значение в рыбоводстве при сортировке рыб по половому признаку.Например, у лососей признаком самцов являются более яркая окраска и изменения опорно-двигательного аппарата - искривление челюстей, появление горба.
У самца колюшки перед нерестом брюшко окрашивается в алый цвет. Самцы многих морских бычков перед нерестом становятся абсолютно черными.У карповых (вобла, язь) на голове и теле самцов в преднерестовый период появляется "жемчужная сыпь" - роговые образования белесого цвета.Появление брачного наряда у рыб определяется гормональными перестройками в этот период жизни. Значение его специалисты трактуют по-разному. Брачная окраска в нерестовый период может выполнять защитную роль. У лососевых брачный наряд маскирует рыб, делает их менее заметными на галечном грунте в прозрачной воде. В других случаях нерестовая окраска имеет сигнальное значение (горбуша, кета). Пол одних рыб можно определить по форме анального отверстия, других - по форме плавников (рис. 1).
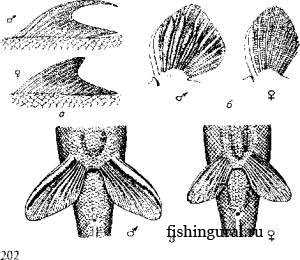 Рис. 1. Вторичные половые признаки рыб:а-спинные плавники Lafaeo dero; б-грудные плавники тибетского гольца; в - брюшные плавники линя
Рис. 1. Вторичные половые признаки рыб:а-спинные плавники Lafaeo dero; б-грудные плавники тибетского гольца; в - брюшные плавники линя Например, у живородящих пициллид половой диморфизм довольно рано проявляется в особом строении анального плавника самцов, выполняющего роль совокупительного органа. У акул такую же роль выполняет придаток брюшного плавника. Важным половым признаком может служить половое поведение рыб перед нерестом. Это может быть преследование самцом самки (карп, щука и др.) или более сложное поведение - устройство гнезда (лабиринтовые), охрана территории. После нереста у многих видов рыб наблюдаются элементы родительского поведения: вентиляция икры, инкубация икры в ротовой полости, охрана молоди (тиляпия).
Конечно, какими бы сложными формами поведения рыбы не обладали в период размножения, все они являются результатом врожденных стереотипических поведенческих актов. Половое поведение проявляется при изменении гормонального статуса рыб в ответ на действие внешней среды. Половая доминанта формируется под влиянием усиления функции гипоталамо-гипофизарно-гонадо-адреналовой системы. Управление половым детерминизмом при искусственном разведении рыб имеет большое практическое значение. Особенно полезным этот прием может быть при разведении ценных рыб - осетровых, лососевых. Здесь желательно иметь большое поголовье самок и ограниченное количество самцов, однако половые признаки у этих видов проявляются довольно поздно, когда уже затрачено много средств и времени.
Регулировать пол в искусственных условиях выращивания удается при помощи стероидных гормонов. Андрогены и эстрогены не разрушаются в желудочно-кишечном тракте. Поэтому они добавляются в корма. Включения метилтестостерона в рацион личинок тиляпии в количестве 30-50 мг/кг приводит к тому, что в стаде половозрелых рыб самцы составляют 95-100 %. При добавлении в рацион форели этого же гормона (3 мг/кг) все особи превращались в самцов.При добавлении в рацион гормона эстрадиола (20 мг/кг корма) у лососей формировалось полностью (на 100 %) женское гомосексуальное стадо. Такой же "феминистический" эффект получали у тиляпии при помощи этинилэстрадиола в количестве 50 мг/кг корма. Таким образом, гормональная регуляция пола у рыб при помощи синтетических или природных стероидов может претендовать на самостоятельный технологический прием в рыбоводстве.
www.fishingural.ru
Есть ли пол у рыб — Все рецепты
Комментарии пользователей Александр Нарышкин 05.11.2017 - 08:23Мороженый кальмар на мелкую терку!
Лена Паскевич 06.11.2017 - 19:41Вторая фотка была разбора галактика
Эдуард Ан 09.11.2017 - 11:23А лекарство не какие. Не капаете?
TheAutoCrash подборка аварий и дтп 10.11.2017 - 16:29она не ответит
Маша Яворская 11.11.2017 - 01:41У меня данио флюоресцентный, у одной рыбки увеличился живот, и и на конце живота тёмное пятнышко, что это значит?
Александр захаренков 13.11.2017 - 20:52Добрый день, такая проблемка начали отдавать желтезной плавники и тело у данио,рыбка активная плавает питается и тд, о чем это говорит?
Людмила Маркина 16.11.2017 - 07:35Спасибо, самое толковое видео
МультДетям 18.11.2017 - 20:55Здравствуйте.где можно купить этих рыбок,сайт не работает
Константин Колмаков 20.11.2017 - 05:05МультДетям в зоомагазине
Александр Иванович 21.11.2017 - 13:56Мне все понятно!
Александр Иванович 24.11.2017 - 11:27Очень приятный голос,😊 и полезное видео👍
Мои аквариумы и их обитатели 26.11.2017 - 07:34Раньше у меня были только розовые данио, после появления данио глофиш у меня стал радужный аквариум. Есть данио всех цветов
Guest 28.11.2017 - 06:00Сасибо за полезное видео! У меня данио и проблема в том,что откусали всем гуппи хвосты и гуппешки умерли,остались данио
Жозе Жозе 01.12.2017 - 01:30Это худшая рыба в аквариуме что у меня была. Несоветую их брать
Natasa Iljina 02.12.2017 - 21:05Сегодня от неристились у меня розовые данио и икры очень много.Подскажите чем ещё можно их кормить как только поплывут?Жидкого корма для мальков у нас в зоомагазине нет и не бывает.Что делать?Инфузории у меня тоже нет. У меня от неристилась гурами и я мальков кормила яичным желтком первое время, а теперь кушают артемию всё хорошо.
Sonya Vern 05.12.2017 - 18:36Снимите пожалуйста о Молинезиях!
Алексей Витион 07.12.2017 - 10:37у меня 1 петушок и 1 данио
Болтай Шерикбай 09.12.2017 - 06:37супер
Mizuki Wolf s Rider 11.12.2017 - 07:45У меня 1 Данио
Bogdan S 12.12.2017 - 17:32а если самка чуток икры метнула, но здоровый живот ещё остался - сколько времени ещё можно их держать в нерестовике?
Bogdan S 14.12.2017 - 02:18А можно садить к красной самке синих самцов? Или наоборот? Ну, чтобы самка и самцы были разного окраса?
Какой то Аноним 15.12.2017 - 16:16Bogdan S. Да)
Хобби TV 15.12.2017 - 13:07а правда што данио рерио пачти халодноводная рибка может жити в температуре16градусах
alena-suveniry.ru
существуют ли рыбы, которые в течение жизни меняют свой пол (с мужского на женский и наоборот).
Представители целого ряда видов рыб могут выступать сразу в двух ипостасях, являясь гермафродитами, или менять свой пол в течение жизни, поочередно переживая радости и превратности как женской, так и мужской судьбы. Такое явление, получившее название последовательного гермафродитизма, сейчас известно для более чем 350 видов рыб, большинство из которых обитают на коралловых рифах. Это представители семейств губановых (Labridae), рыб-попугаев (Scaridae), групперов (Serranidae), помацентровых (Pomacentridae) и некоторых других. Самец шишкоголовой рыбы-попугая (Bolbometopos muricatum) У многих из них все мальки, вылупляющиеся из икринок, – самки. Вырастая, они созревают, один или несколько раз откладывают икру, а затем превращаются в самцов и вновь принимают участие в размножении. Такая форма последовательного гермафродитизма называется протогинией. Впрочем, у некоторых видов рыб-попугаев* и губанов часть мальков рождается все же самцами. В отличие от своих сестер – будущих братьев – они остаются верны своему полу всю жизнь. Правда, в зрелом возрасте этих рыб – первичных самцов – трудно отличить от тех, кто провел молодость в качестве представительниц прекрасного пола. И первичные, и вторичные самцы во второй половине своей жизни не только выглядят сходно, но и придерживаются одинаковой жизненной стратегии, подходящей остепенившимся и солидным индивидам. Вместе с крупными размерами, делающими их неуязвимыми для многих мелких хищников, они приобретают яркую окраску – голубую, зеленую, красную или оранжевую, а в ряде случаев – и горбовидные выросты на лбу. Недаром взрослые рыбы-попугаи считаются одними из самых красивых обитателей коралловых садов! Однако внешний лоск – не самое главное. Дожившие до зрелого возраста рыбы обзаводятся и кое-чем посущественнее – собственным участком на рифе, который активно охраняют от вторжения других самцов и куда привлекают самок. Правда, размеры семенников и, соответственно, количество продуцируемых молок у этих самцов не особенно велики, но им, как говорится, хватает. Ведь самка, привлеченная возможностью проводить время на безопасном и кормном участке, отложит икру в нужное время и в нужном месте, так что даже небольшого количества молок хватит, чтобы надежно оплодотворить кладку. К тому же, несмотря на кажущуюся умеренность, зрелые рыбы-попугаи регулярно меняют подруг и на протяжении репродуктивного сезона умудряются облагодетельствовать немало охочих до респектабельной жизни самок. Причем значительно более юного, чем они сами, возраста. А как же иначе? Ведь «старух» у протогинических рыб просто не бывает!
про рыб не знаю..а креветки точно меняют...и не один раз....это зависит от температуры воды..если теплые воды..то чаще....=)
Точно слышала, что несколько видов есть, и даже более высших животных (лягушки). Но вот названия не вспомню, надо искать.
да,существуют. не помню названия,но их особенность в том,что меняют они пол только при отсутствии противоположного. позже могут и обратно поменять.
Существуют... это меченосцы!
Аквариумные меченосцы меняют свой пол по мере надобности.
Видел сам как в школьном аквариуме в плохих условиях меченоска сменила пол на мужской: посадили пару, а потом стало два самца.
Конечно могу ошибаться, но меченостцы меняют пол только с самки на самца. В обратную сторону не сталкивался.
Гермафродитизм у рыб - явление редкое, но не уникальное, - Чаще это бывает у каспийской сельди. Такие рыбы вполне здоровы и даже способны метать икру. Впрочем, с полной уверенностью утверждать, что эта рыба - именно гермафродит, нельзя. Дело в том, что пол у рыб не жестко запрограммирован и может меняться в течение жизни. На стадии малька у многих рыб нет пола, они становятся самками или самцами в зависимости от того, что требуется данной популяции. Пол может поменяться и у взрослой рыбы, даже после того, как она уже метала икру. Иногда рыба может становиться то самкой, то самцом несколько раз. От чего зависит перемена пола, точно неизвестно. Замечено, что пол рыб меняется в зависимости от солености воды, ее температуры, корма, глубины, на которой плавает косяк.
это рыба- Клоун, губан обитающий в Красном море
touch.otvet.mail.ru
Смена пола у рыб
В естественных условиях смена пола это довольно частое явление среди морских рыб, у которых преобладает трансформация самцов в самок [8; 13; 21; 27]. В данном случае пол определяется рядом генетических факторов, а также факторами среды, например, pH и температурой. После наступления половозрелости поведенческие взаимодействия могут запустить начало смены пола, то есть наблюдается последовательный гермафродитизм. В большинстве случаев особи, получившие во время взросления один пол, затем необратимо меняют его на противоположный. У протогенных видов особи трансформируются от самок в самцов, у протандрических — первостепенным полом является мужской, который затем сменяется некоторыми рыбками на женский.
Одна из гипотез, объясняющая целесообразность гермафродитизма, в качестве ключевого аспекта рассматривает размер тела. Согласно ей, рыбки меняют пол в зависимости от ожидаемой прибавки в размерах. Как правило, плодовитость самцов, в частности, количество продуцируемых гамет, не зависит от их длины и массы. То есть самцы любой длины имеют одинаковую плодовитость. Икринки значительно крупнее сперматозоидов, отсюда, чем больше самка, тем она более плодовита. Будучи маленьким и молодым, самец в полной мере проявляет свой репродуктивный потенциал, но достигнув критического размера превращается в самку (протандрический вид). У протогенных рыбок трансформация пола призвана максимизировать размеры самцов, что объясняется важностью охраны гарема и своей территории [28].
У некоторых гонохористичных (гонохоризм — раздельнополость) рыбок, у которых пол четко дифференцирован на протяжении всей жизни, половые отличия начинают проявляться уже на ранней стадии развития. Однако среди гонохор имеются представители, являющиеся самками на раннем этапе онтогенеза, а затем при посредничестве генетических факторов, трансформирующиеся в самцов [25]. Часто после изменения пола в гонадах у самцов данных видов остаются ооциты.
Изначальная представленность популяции рудиментарных гермафродитов только самками навела на мысль, что женский пол является полом "по-умолчанию" [22]. Даже протандрические рыбки изначально не дифференцируются как самцы. В частности, гонады неполовозрелого Клоуна меланопуса (Amphiprion melanopus — Pomacentridae) содержат преимущественно женскую ткань. Тестикулярные элементы начинают преобладать во время созревания особей в самцов, однако они исчезают при трасформации в самку [10; 22]. Подобный механизм был обнаружен и у других видов рода Amphiprion и у аквариумных экземпляров Sparus aurata (Sparidae) [22]. Кроме того, у молоди протандрических видов семейства Creediidae обнаружены межполовые гонады, позднее созревающие преимущественно в мужские [14]. Таким образом, мужской пол протадрических видов является скорее переходной, нежели начальной стадией трансформации.
Хотя гермафродитизм обычен для рифовых морских рыб, он крайне редко встречается у пресноводных видов. В природе естественная смена пола последними плохо изучена, поэтому причисление некоторых из них к животным с дихогамией (последовательный гермафродитизм) является скорее исключением из правил. Одним из первых, документально зафиксированным случаем смены пола в естественной среде обитания можно назвать транформацию особей вида Monopterus albus, рисового угря [5; 6].
Среди пресноводных обитателей самыми исследуемыми в вопросе половой дифференцировки являются Цихлиды. Это связано с чрезвычайным разнообразием стратегий размножения, включающих откладывание икры на субстрат, либо инкубацию во рту [2], опеку за потомством одним [9], обоими родителями [1], групповую [23; 26] или даже межвидовую [16] заботу за молодью, сложную систему брачных связей (моногамия, полигамия, стратегия ухода).
Множество работ описывают различные проявления половой лабильности данного семейства пресноводных рыбок. Одним из первых является исследование Полдера, который в 1971 году на примере Aequidens portalegrensis открыл феномен смены пола у южно-американских цихлид [18]. Это породило гипотезу о возможности смены пола у других цихловых. Она была подкреплена Питерсом (1975) [19], который при проведении гистологических исследований обнаружил в гонадах самцов 14 видов малавийских цихлид как ооциты, так и сперматогонии. Данные экспериментов свидетельствуют о том, что у изученных представителей группы Мбуны наблюдается гермафродитизм, чаще проявляющийся в трансформации самца в самку. Позднее, Запанс (1985) [29] заявил, что особи вида Cynotilapia afra, члена группы Мбуна, также изменяют пол от самца к самке. Однако данное исследование не включает специальной проверки заявляения.
Полевые наблюдения Рибинка и коллег (1983) [20] продемонстрировали, что все члены 200 близких видов группы Мбуна озера Малави вынашивают икру во рту. Но среди некоторых представителей Pseudotropheus lombardoi (Барджесс, 1977) [3] вынашивают икру особи, имеющие окраску самцов. Имеется неколько возможных объяснений этого феномена, включающие вынашивание икры взрослым самцом, самкой, имеющей как самцовую, так и обычную окраску, либо смену пола без сопутствующих изменений окраски.
При проведении поведенческих исследований с последующим анатомическим вскрытием [17]. было показано, что в семенниках всех самцов находятся недоразвитые ооциты. Это говорит о прохождении гонад Pseudotropheus lombardoi через интерсекуальный ювенильный период. На поздней стадии развития рыбок, тестикулярные элементы доминируют в гонадах, хотя последние и содержат незрелую ткань ооцитов. Самки Pseudotropheus lombardoi проявляют диморфизм, когда некоторые из них имеют сине-желтую окраску, а другие традиционную синюю. Кроме анатомических перестроек у трансформированных самок наблюдаются поведенческие реакции, аналогичные самцам, например, охрана территории и строительство гнезда. Половая система трансформированных самок не притерпевает изменений, в гонадах присутствуют ооциты на различных стадиях развития, отсутствует атрезия ооцитов и развитие ткани семенников. Таким образом, самки не утрачивают детородную функцию, но при прочих равных, они пользуются гораздо меньшим предпочтением со стороны самцов по сравнению с обычными самками.
Более сложная картина транформации наблюдалась при изучении малавийского вида Metriaclima cf. livingstoni [12]. В эксперименте самца отделяли от трех самок с помощью прозрачного пластика. После удаления самца, у одной из самок развивались самцовые вторичные половые признаки, и она начинала оплодотворять икру двух других особей. Во втором эксперименте 6-7 функциональных самок помещалось в аквариум, и в отсутствии самца вновь происходила смена пола самки, которая приступала к нересту и оплодотворяла икру.
Аналогичные результаты получены для вида Crenicara punctulata, который по результатам поведенческих исследований и изучению гистологии половой системы трансформированных особей признан протогенным. [4].
Таким образом, при рассмотрении разнообразия цихлид можно отметить присутствие у некоторых видов предрасположенности к смене пола (фенотипической или полной). Малая распространенность последовательного гермафродитизма у данного семейства и возможно других пресноводных рыб, вероятно, объясняется невозможностью прогнозирования присутствия пищи и полового партнера по сравнению с коралловыми рыбами. Так, у морских рыб очень распространено поддержание постоянных мест кормления, которые особи охраняют. По наблюдениям Гранта (1997) постоянные кормовые угодья наблюдаются у 67% морских коралловых рыб и лишь у 6% морских рыб вод восточной Канады и 9% пресноводных рыб Канады [11]. Благодаря изобилию особей-потенциальных половых партнеров в местах кормления, пища формирует социальную структуру [7]. Тем не менее, различия в защите кормовых угодий не настолько сильные: 84% коралловых рифовых рыбок охраняют эти области, а также места нереста и свое потомство; тоже самое делают 73% морских рыб восточной Канады и 68% пресноводных [11]. Кроме того, спаривание на коралловых рифах обычно происходят продолжительное время [24], тогда как тропические пресноводные рыбки нерестятся сезонно [15].
Короткие периоды размножения, обычные для озерных и речных рыб, а также отсутствие постоянных областей кормления не позволяют предсказывать распределение половых партнеров. Однако гипотеза, включающая размерную модель (см. выше), нуждается в точной оценке будущего репродуктивного потенциала. То есть основой для формирования последовательного гермофродитизма служит определенность (места и времени) или прогнозируемость предстоящего спаривания.——Использованные источники:Oldfield R.G. Genetic, abiotic and social influences on sex differentiation in cichlid fishes and the evolution of sequential hermaphroditism. Fish and Fisheries, 2005, 6, 93–110
1. Barlow G.W. (1976) The Midas cichlid in Nicaragua. In: Investigations of the Ichthyofauna of Nicaraguan Lakes (ed. T.B. Thorson). School of Life Sciences, University of Nebraska, Lincoln, Nebraska, pp. 333–358.2. Barlow G.W. (2001) Parsing mouth brooding in cichlid fishes to illuminate the course of evolution. Journal of Aquariculture and Aquatic Sciences, Cichlid Research: State of the Art 9, 66–78.3. Burgess W. E. 1977. Pseudotropheus lombardoi, a new species of Lake Malawi Mbuna with reversed sexual coloration (Pisces: Cichlidae) . T.F.H . Magazine 26 : 63-67.4. Carruth L.L. Freshwater Cichlid Crenicara punctulata Is a Protogynous Sequential Hermaphrodite. Copeia 2000(1):71-82. 20005. Chan S. T. H., Philips J. G. (1967a). The structure of the gonad during natural sex reversal in Monopterus albus (Pisces, Teleostei). J. Zool., Lond. 151: 169-1416. Chan S. T. H., Philips J. G. (1967b). Seasonal changes in the distribution of gonadal lipids and spermatogonic tissue in the male phase of Monopterus albus (Pisces, Teleostei). J. Zool.. Lond. 152: 31-417. Emlen, S.T. and Oring, L.W. (1977) Ecology, sexual selection, and the evolution of mating systems. Science 197, 215–223.8. Francis R. C., 1992. Sexual lability in teleosts: developmental factors. Q Rev Biol 67:1-189. Fryer G. and Iles T.D. (1972) The Cichlid Fishes of the Great Lakes of Africa: Their Biology and Evolution. Oliver and Boyd, Edinburgh, UK.10. Godwin J.R. (1994) Histological aspects of protandrous sex change in the anemonefish Amphiprion melanopus. Journal of Zoology (London) 232, 199–213.11. Grant J.W.A. (1997) Territoriality. In: Behavioural Ecology of Teleost Fishes (ed. J.-G.J. Godin ), Oxford University Press, Oxford, pp. 81–103.12. Jay R. Stauffer Jr and Renea A. Ruffing and (2008) Behaviorally Induced Sex Reversal of Metriaclima cf. livingstoni (Cichlidae) from Lake Malawi. Copeia: September 2008, Vol. 2008, No. 3, pp. 618-620.13. Kuwamura T., Nakashima Y., 1998. New aspects of sex change among reef fishes: recent studies from Japan. Environ Biol Fishes 52:125-13514. Langston R.C. (2004) Gonad Morphology and Sex Change in Sandburrowers (Teleostei: Creediidae). PhD Dissertation, University of Hawaii, Manoa, 138 pp.15. Lowe-McConnell R.H. (1975) Fish Communities in Tropical Freshwaters: Their Distribution, Ecology, and Evolution. Longman, London, 337 pp.16. McKaye K.R. (1977) Defense of a predator’s young by a herbivorous fish: an unusual strategy. American Naturalist 111, 301–315.17. Naish K., Ribbink A. J. A preliminary investigation of sex change in Pseudotropheus lombardoi // Environmental Biology of Fishes 28: 285 -294, 1990.18. Polder J. J. W. 1971. On gonads and reproductive behaviour in the cichlid fish Aequidens portalegrensis (Hensel). Neth. J. Zool. 21: 265-365.19. Peters H. M. 1975. Hermaphroditism in cichlid fishes . pp . 228-235. In: R. Reinboth (ed.) Intersexuality in the Animal Kingdom. Springer-Verlag, Berlin.20. Ribbink A. J., Marsh B. A., Marsh A. C., Ribbink A. C., Sharp B. J. 1983. A preliminary survey of the cichlid fishes of rocky habitats in Lake Malawi. S. Afr. J. Zool. 18 : 149-310.21. Shapiro D. Y., 1987. Sexual differentiation, social behavior, and the evolution of sex change in coral reef fishes. Biosci 37:490-49722. Shapiro D.Y. (1992) Plasticity of gonadal development and protandry in fishes. Journal of Experimental Zoology 261, 194–203.23. Taborski M. (1984) Broodcare helpers in Lamprologus brichardi: their costs and benefits. Animal Behaviour 32, 1236–1252.24. Thresher R.E. (1984) Reproduction in Reef Fishes. T.F.H. Publications, Neptune City, NJ. 399 pp.25. Uchida D., Yamashita M., Kitano T.and Iguchi T.(2002) Oocyte apoptosis during the transition from ovary-like tissue to testes during sex differentiation of juvenile zebrafish. Journal of Experimental Biology 205, 711–718.26. Ward J.A. and Wyman R.L. (1977) Ethology and ecology of cichlid fishes of the genus Etroplus in Sri Lanka: preliminary findings. Environmental Biology of Fishes 2, 137–145.27. Warner R. R., 1984. Mating behavior and hermaphroditism in coral reef fishes. Am Sci 72:128-13628. Warner R. R. (1975) The adaptive significance of sequential hermaphroditism in animals. The American Naturalist 109, 61–8229. Zupanc, G .K .H . 1985. Fish and their behavior . Tetra Verlag, der Gattungen Tropheus, Petrochromis, Haplochromis and Melle. 188 pp.
aquavitro.org
есть ли у рыб пенис?
У живородящих рыб часть анального плавника модернизировалась в орган, имеющий форму трубки. который называется гоноподиумом. он используется для оплодотворения самок. У икромечущих рыб представителей разных полов различить гораздо труднее.
жаль что рыбы не разговоаривают, они бы тебе точно ответили по полной...)))
ннезнаю не проверяла=)))!
У акуловых рыб анальные плавники тоже преоразованы в подобие пениса. Но в общем-то им это без надобности - оплодотворение наружное.
Бывает, и ещё какой <a rel="nofollow" href="http://ekabu.ru/animals/21742-chudesa-prirody-ryba-chlen-guidak.html" target="_blank">http://ekabu.ru/animals/21742-chudesa-prirody-ryba-chlen-guidak.html</a>
touch.otvet.mail.ru














.png)
.png)
.png)