Какова симметрия тела сцифоидных медуз. Как устроены основные системы органов сцифоидных. Какое эволюционное значение имеет симметрия тела сцифоидных медуз. Как происходит жизненный цикл сцифоидных.
Содержание
Симметрия тела сцифоидных медуз
Сцифоидные медузы относятся к типу Кишечнополостных (Cnidaria) и классу Сцифоидных (Scyphozoa). Их тело имеет радиальную (лучевую) симметрию, в основе которой лежит четырехлучевой план строения. Что это означает?
Основные особенности симметрии сцифоидных медуз:
- Тело имеет форму колокола или зонтика
- Через центр тела проходит главная ось симметрии
- От центра отходят 4 радиальных канала, расположенных крестообразно
- Внутренние органы и структуры повторяются 4 раза вокруг главной оси
- Край зонтика разделен на 8 краевых лопастей
Таким образом, через тело сцифоидной медузы можно провести 4 плоскости симметрии. Это определяет радиально-симметричный план строения, характерный для большинства кишечнополостных.

Эволюционное значение радиальной симметрии
Радиальная симметрия тела сцифоидных медуз имеет важное эволюционное значение:
- Позволяет воспринимать раздражения со всех сторон одинаково
- Обеспечивает равномерное распределение щупалец по краю тела
- Способствует активному плаванию в толще воды
- Является адаптацией к пассивно-плавающему образу жизни
Однако радиальная симметрия ограничивает возможности активного движения в определенном направлении. Поэтому у более высокоорганизованных животных в ходе эволюции возникла двусторонняя симметрия тела.
Особенности внутреннего строения сцифоидных медуз
Несмотря на внешнюю радиальную симметрию, во внутреннем строении сцифоидных медуз можно обнаружить элементы двусторонней симметрии. Рассмотрим основные системы органов:
Пищеварительная система
Пищеварительная система сцифоидных медуз включает следующие элементы:
- Рот, расположенный в центре нижней стороны зонтика
- Короткая трубчатая глотка
- Желудок, от которого отходят радиальные каналы
- Кольцевой канал по краю зонтика
В желудке имеются 4 гастральные нити, выделяющие пищеварительные ферменты. Это создает элемент четырехлучевой симметрии.

Нервная система
Нервная система сцифоидных диффузного типа, но имеет некоторую централизацию:
- Нервное кольцо по краю зонтика
- Скопления нервных клеток в ропалиях (органах чувств)
- 8 краевых ганглиев, связанных с ропалиями
Наличие парных ганглиев и органов чувств создает элементы билатеральной симметрии в нервной системе.
Органы чувств
По краю зонтика медузы расположены специализированные органы чувств — ропалии. Их обычно 8 штук, что соответствует числу краевых лопастей. В каждом ропалии имеются:
- Статоцист (орган равновесия)
- Светочувствительные глазки
- Химические рецепторы
Парное расположение ропалиев также создает элемент двусторонней симметрии.
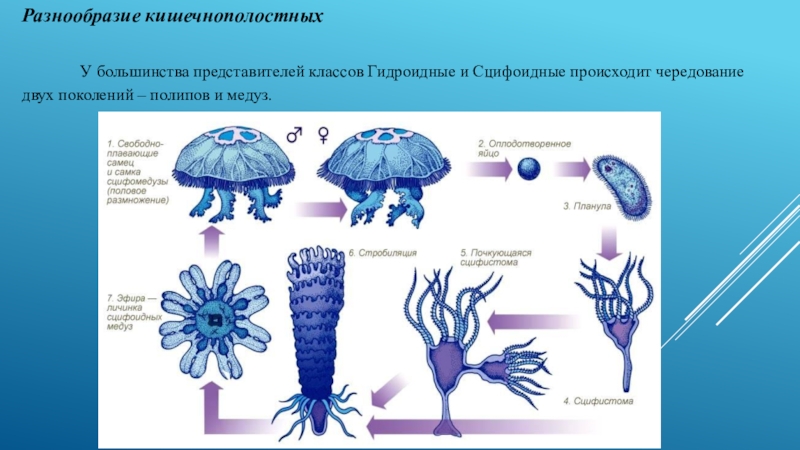
Жизненный цикл сцифоидных медуз
Жизненный цикл сцифоидных медуз включает чередование полового и бесполого поколений:
Основные стадии:
- Половая стадия — медуза
- Планула — свободноплавающая личинка
- Сцифистома — прикрепленный полип
- Стробила — почкующийся полип
- Эфира — молодая медуза
Интересно, что полипоидная стадия (сцифистома) имеет радиальную симметрию, как и взрослая медуза. Однако в процессе стробиляции и образования эфир проявляются элементы двусторонней симметрии.
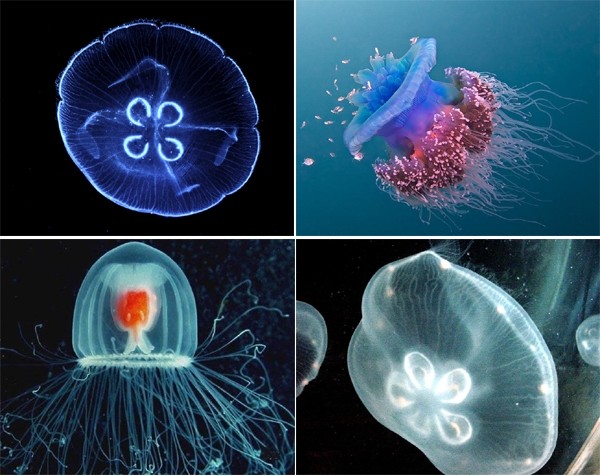
Как происходит образование медуз?
Процесс образования молодых медуз называется стробиляцией и включает следующие этапы:
- На теле сцифистомы образуются поперечные перетяжки
- Верхняя часть тела делится на несколько дисков
- Каждый диск развивается в молодую медузу — эфиру
- Эфиры отрываются и уплывают
- Из оставшейся нижней части сцифистомы регенерирует новый полип
Таким образом, в жизненном цикле сцифоидных медуз чередуются стадии с радиальной и двусторонней симметрией, что отражает их эволюционную историю.
Эволюционное происхождение симметрии тела медуз
Вопрос о происхождении симметрии тела кишечнополостных вызывает споры среди ученых. Существуют две основные гипотезы:
Гипотеза первичности радиальной симметрии
Согласно традиционным представлениям:
- Радиальная симметрия кишечнополостных первична
- Двусторонняя симметрия билатерий возникла позже
- Общий предок имел радиально-симметричное строение
Гипотеза первичности двусторонней симметрии
Новые данные свидетельствуют в пользу другой гипотезы:
- Двусторонняя симметрия была исходной для кишечнополостных
- Радиальная симметрия медуз — вторичное приобретение
- Общий предок имел билатерально-симметричное строение
В пользу второй гипотезы говорят следующие факты:
- Наличие элементов двусторонней симметрии у кораллов
- Билатеральное расположение зачатков органов у личинок
- Сходство экспрессии генов развития с билатериями
Таким образом, симметрия тела сцифоидных медуз отражает сложную эволюционную историю кишечнополостных животных. Сочетание радиальной и двусторонней симметрии позволяет им эффективно существовать в водной среде.
Сцифоидные | это… Что такое Сцифоидные?
Сцифо́идные (сцифомеду́зы, лат. Scyphozoa) — класс морских организмов из типа стрекающих (Cnidaria). Группа включает сравнительно небольшое количество видов — около 200. Жизненный цикл сцифоидных, как правило, представляет собой метагенез, в котором присутствует бесполая (полипоидная) и половая (медузоидная ) стадии. Медузы некоторых представителей характеризуются крупными размерами и порой образуют очень крупные скопления. Полипы сцифоидных (сцифистомы), напротив, обладают очень мелкими размерами — порядка нескольких миллиметров.
Обычные для морей России сцифоидные — ушастая медуза (Aurelia aurita), львиная грива (Cyanea capillata), корнерот (Rhizostoma pulmo).
Содержание
|
Общий план строения
Диаметр зонтика обычно составляет от 2 до 40 см, но встречаются и ещё более крупные медузы, например Cyanea capillata, достигающая 2 м в поперечнике. Внешний вид сцифоидных медуз зачастую весьма необычен, особенно когда гонады и другие внутренние органы, ярко окрашенные в тёмно-оранжевый, розовый или иные насыщенные цвета, просвечивают сквозь прозрачный или слегка окрашенный колокол.
Внешний вид сцифоидных медуз зачастую весьма необычен, особенно когда гонады и другие внутренние органы, ярко окрашенные в тёмно-оранжевый, розовый или иные насыщенные цвета, просвечивают сквозь прозрачный или слегка окрашенный колокол.
Как и у всех кишечнополостных, тело сцифоидных состоит из двух эпителиальных пластов: выстилки поверхности тела (эпидермиса) и выстилки кишечной полости (гастродермиса). Между ними находится мезоглея — желеобразная соединительная ткань, на 98 % состоящая из воды, содержащая белки и гликопротеиды (коллаген, ламинин, фибронектин). Кроме того, в мезоглее находится сравнительно небольшое количество клеток, мигрировавших из эпителиальных пластов.
В зависимости от вида сцифистомы могут быть одиночными или колониальными. Тело полипа, расширенное на оральном конце, резко сужается примерно посередине. От середины до педального диска тело полипа совсем узкое. В основе строения сцифоидных лежит радиальная (четырёхлучевая) симметрия. Ось этой симметрии проходит между ротовым отверстием и подошвой (у полипов) или вершиной зонтика (у медуз). Симметрия выражается в наличии четырёх септ в гастральной полости. Они равноудалены друг от друга и разделяют гастральную полость на четыре части. Септы имеют наибольшую ширину в месте прикрепления к оральному диску, сужаются в аборальном направлении и совсем исчезают, немного не доходя до педального конца полипа. В отличие от коралловых полипов септы сцифистом не несут мезентериальных нитей, но в каждой септе имеется септальная (субумреллярная) воронка, образующаяся в результате инвагинации орального диска. Начинаясь на поверхности орального диска, воронка заходит глубоко в толщу септы, где слепо заканчивается. Эпителиально-мускульные клетки, выстилающие каждую из четырёх воронок, образуют продольные мышцы-ретракторы. Полюс тела, на котором расположен рот, называют оральным, противоположный ему — аборальным, а ось симметрии, соответственно, — орально-аборальной.
Ось этой симметрии проходит между ротовым отверстием и подошвой (у полипов) или вершиной зонтика (у медуз). Симметрия выражается в наличии четырёх септ в гастральной полости. Они равноудалены друг от друга и разделяют гастральную полость на четыре части. Септы имеют наибольшую ширину в месте прикрепления к оральному диску, сужаются в аборальном направлении и совсем исчезают, немного не доходя до педального конца полипа. В отличие от коралловых полипов септы сцифистом не несут мезентериальных нитей, но в каждой септе имеется септальная (субумреллярная) воронка, образующаяся в результате инвагинации орального диска. Начинаясь на поверхности орального диска, воронка заходит глубоко в толщу септы, где слепо заканчивается. Эпителиально-мускульные клетки, выстилающие каждую из четырёх воронок, образуют продольные мышцы-ретракторы. Полюс тела, на котором расположен рот, называют оральным, противоположный ему — аборальным, а ось симметрии, соответственно, — орально-аборальной. Четырёхлучевая симметрия хорошо прослеживается в строении специализированного манубриума. Он может быть квадратным в сечении или же удлинён и разделён на четыре ротовые лопасти, которые тянутся за зонтиком плывущей медузы. Щупальца, расположенные по краю зонтика, иногда собраны в четыре или восемь групп, но часто они просто равномерно распределены по его периметру. Они прикрепляются к субстрату хитиновой перидермой. Перидерма может покрывать или только нижнюю часть, или всё тело полипа.
Четырёхлучевая симметрия хорошо прослеживается в строении специализированного манубриума. Он может быть квадратным в сечении или же удлинён и разделён на четыре ротовые лопасти, которые тянутся за зонтиком плывущей медузы. Щупальца, расположенные по краю зонтика, иногда собраны в четыре или восемь групп, но часто они просто равномерно распределены по его периметру. Они прикрепляются к субстрату хитиновой перидермой. Перидерма может покрывать или только нижнюю часть, или всё тело полипа.
Пищеварительная система
Вокруг рта находится поле, обрамлённое венчиком щупалец, — оральный диск. Четыре септы, расположенные на одинаковом расстоянии друг от друга, делят пищеварительную полость на центральный желудок и четыре желудочных кармана. Толстые септы, сохранившиеся от полипа в почти неизменном виде, содержат открывающиеся на оральную поверхность септальные воронки. В непосредственной близости от них находятся четыре пары гонад, которые, как и у коралловых полипов, залегают на поверхности септ. В воронках создаётся, по всей видимости ресничками, ток воды. Возможно, благодаря этому гонады снабжаются кислородом. Свободные края септ несут пучки тонких, нитевидных гастральных нитей. Свободный конец каждой нити свешивается в желудок. Гастральные нити напоминают книдожелезистые тяжи и аконции представителей Anthozoa. Они несут нематоцисты и выделяющие ферменты железистые клетки и играют важную роль в обездвиживании и переваривании добычи. У некоторых сцифоидных медуз от желудка отходят радиальные каналы. Неоднократно ветвясь, они образуют в толстом слое мезоглеи гастроваскулярную систему. Радиальные каналы часто соединяются с кольцевым каналом, проходящим по краю зонтика. Реснички гастродермиса прогоняют содержащуюся в гастроваскулярной системе жидкость по определённому пути. Если желудок можно считать «сердцем» этой системы, то одни радиальные каналы — «артерии», уносящие жидкость от желудка, а другие — «вены», приносящие жидкость обратно в желудок. Толстый слой мезоглеи содержит амебоциты, которые, возможно, несут функцию фибробластов.
В воронках создаётся, по всей видимости ресничками, ток воды. Возможно, благодаря этому гонады снабжаются кислородом. Свободные края септ несут пучки тонких, нитевидных гастральных нитей. Свободный конец каждой нити свешивается в желудок. Гастральные нити напоминают книдожелезистые тяжи и аконции представителей Anthozoa. Они несут нематоцисты и выделяющие ферменты железистые клетки и играют важную роль в обездвиживании и переваривании добычи. У некоторых сцифоидных медуз от желудка отходят радиальные каналы. Неоднократно ветвясь, они образуют в толстом слое мезоглеи гастроваскулярную систему. Радиальные каналы часто соединяются с кольцевым каналом, проходящим по краю зонтика. Реснички гастродермиса прогоняют содержащуюся в гастроваскулярной системе жидкость по определённому пути. Если желудок можно считать «сердцем» этой системы, то одни радиальные каналы — «артерии», уносящие жидкость от желудка, а другие — «вены», приносящие жидкость обратно в желудок. Толстый слой мезоглеи содержит амебоциты, которые, возможно, несут функцию фибробластов.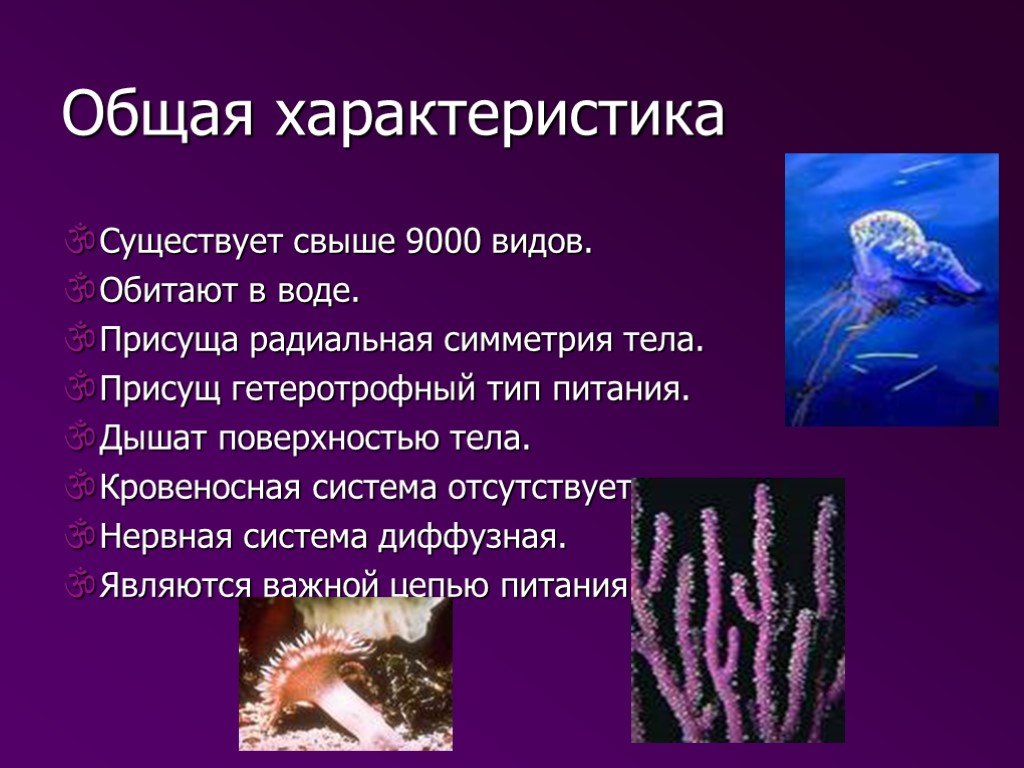 Кишечник сцифоидных слепо замкнут, так что ротовое отверстие поочерёдно выполняет функции рта и ануса. Полипы Scyphozoa в отличие от коралловых полипов лишены глотки и орального сфинктера.
Кишечник сцифоидных слепо замкнут, так что ротовое отверстие поочерёдно выполняет функции рта и ануса. Полипы Scyphozoa в отличие от коралловых полипов лишены глотки и орального сфинктера.
Органы чувств
Край тела медузы часто фестончатый и подразделён на широкие, плавно закруглённые лопасти (лаппеты), которые способствуют изгибанию сокращающегося зонтика. По краю последнего, на равных расстояниях друг от друга, в выемках между лопастями расположены органы чувств — ропалии. Ропалии — настоящие органы; они образованы производными гастродермы, мезоглеи и эпидермы. Сенсорные структуры ропалия включают статоцист гастродермального происхождения и механорецептор, вероятно, также хеморецептор, а иногда ещё и фоторецептор. Хотя по сравнению с органами чувств человека ропалии кажутся примитивными, они успешно обеспечивают медузу разнообразной информацией. Ропалии регистрируют направление силы тяжести (чувство равновесия), распространяющиеся в воде колебания (слух), химические сигналы (запах) и положение источника света. Полученная сенсорная информация поступает в расположенный у основания ропалия ганглий. Здесь она обрабатывается, и соответствующие сигналы передаются в нервную сеть и нервное кольцо. Ганглии, расположенные у основания ропалиев, имеют пейсмейкерные нейроны, на частоту импульсов которых влияет полученная от ропалия информация. За счёт этого осуществляется контроль над скоростью и направлением плавания. Субумбреллярные локомоторные мышцы представлены поперечнополосатой кольцевой (корональной) мышцей и радиальными мышцами.
Полученная сенсорная информация поступает в расположенный у основания ропалия ганглий. Здесь она обрабатывается, и соответствующие сигналы передаются в нервную сеть и нервное кольцо. Ганглии, расположенные у основания ропалиев, имеют пейсмейкерные нейроны, на частоту импульсов которых влияет полученная от ропалия информация. За счёт этого осуществляется контроль над скоростью и направлением плавания. Субумбреллярные локомоторные мышцы представлены поперечнополосатой кольцевой (корональной) мышцей и радиальными мышцами.
Питание
Взрослые сцифоидные медузы питаются мелкими животными, особенно рачками, а многие поедают также других медуз. Некоторые медузы питаются рыбой, но, с другой стороны, мальки многих рыб держатся рядом с представителями определённых видов сцифоидных медуз, находя у них защиту. Медленно плавая или плавно погружаясь в толщу воды, медуза захватывает добычу, которая контактирует с щупальцами или манубриумом. Щупальца могут подгибаться или сокращаться, подтягивая добычу ближе к манубриуму. У полипов и медуз Schyphozoa нематоцисты залегают в эпидермисе, а у медуз также в гастродермисе (в гастральных нитях).
У полипов и медуз Schyphozoa нематоцисты залегают в эпидермисе, а у медуз также в гастродермисе (в гастральных нитях).
Пищевое поведение медуз представляет собой сложный процесс. Движение питающихся планктоном аурелий получило название «кульбитрующего», так как представляет собой серию повторяющихся «мёртвых петель». Благодаря этому медуза может захватить больший объём воды, чем при движении по прямой. Для «сбора» планктона используется эксубрелла, функционирующая как большая «липучка». Этому способствует наличие в эпидермисе эксумбреллы многочисленных скоплений книдоцитов и секреторных клеток, выделяющих слизь. Слизистые выделения с приклеившимися к ним мелкими организмами постепенно перемещаются к краю зонтика. Здесь слизь между основаниями щупалец перетекает на субумбреллярную поверхность и скапливается в специальном узком желобке, проходящем параллельно краю зонтика. Медуза постоянно «облизывает» этот желобок ротовыми лопастями, собирая комочки слизи. Затем слизь с содержащимися в ней пищевыми объектами с помощью ресничных клеток, расположенных на ротовых лопастях, перемещается к ротовому отверстию и поступает в желудок.
Жизненный цикл
Как правило, полипы Scyphozoa (сцифистомы) представляют собой бесполое поколение жизненного цикла, а медузы — половое. Рост колониальных видов и бесполое размножение одиночных форм сцифистом происходит путём почкования. Почки закладываются на теле полипа или, например у Aurelia, на столоне. В соответствующее время года под влиянием гормональных изменений и факторов окружающей среды на сцифистомах бесполым путём образуются молодые медузы. Этот процесс называется стробиляцией. На оральном конце стробилирующей сцифистомы (теперь это уже стробила) (в литературе стробилой, как правило, называют не приступившего к стробиляции сцифополипа, а лишь ещё не отделившихся от него эфир. В зависимости от количества последних обычно говорят о монодисковой или полидисковой стробиле. Сам полип в результате интенсивной полидисковой стробиляции может сильно уменьшаться в размерах, терять щупальца, но, как только стробиляция заканчивается, он довольно быстро восстанавливается) закладывается дисковидная медуза. Позднее она отделяется от стробилы в результате образования поперечной, кольцевой перетяжки. Только что отделившаяся свободно плавающая ювенильная медуза называется эфирой. Эфира имеет небольшой диаметр и сильно изрезанный край зонтика, лопасти которого при плавании совершают энергичные взмахи. В зависимости от вида на сцифистоме образуется одна медуза (монодисковая стробиляция) или много медуз (полидисковая стробиляция). У форм с полидисковой стробиляцией, например у видов рода Aurelia и других распространённых представителей Scyphozoa, совокупность расположенных одна над другой развивающихся эфир напоминают стопку блюдечек. На вершине стробилы находятся эфиры, которые начали формироваться прежде других и которые соответственно отделятся от неё первыми. После завершения стробиляции сцифистома продолжает существовать как самостоятельный полип до следующего года, когда снова начинается процесс стробиляции, и на ней снова образуются эфиры. Сцифистома может жить от одного года до нескольких лет.
Позднее она отделяется от стробилы в результате образования поперечной, кольцевой перетяжки. Только что отделившаяся свободно плавающая ювенильная медуза называется эфирой. Эфира имеет небольшой диаметр и сильно изрезанный край зонтика, лопасти которого при плавании совершают энергичные взмахи. В зависимости от вида на сцифистоме образуется одна медуза (монодисковая стробиляция) или много медуз (полидисковая стробиляция). У форм с полидисковой стробиляцией, например у видов рода Aurelia и других распространённых представителей Scyphozoa, совокупность расположенных одна над другой развивающихся эфир напоминают стопку блюдечек. На вершине стробилы находятся эфиры, которые начали формироваться прежде других и которые соответственно отделятся от неё первыми. После завершения стробиляции сцифистома продолжает существовать как самостоятельный полип до следующего года, когда снова начинается процесс стробиляции, и на ней снова образуются эфиры. Сцифистома может жить от одного года до нескольких лет.
Эфирам некоторых видов нужно два года, чтобы превратиться в половозрелых медуз, в то время как у других видов стадия эфиры относительно непродолжительна. Эфиры Aurelia aurita на западном побережье США образуются в марте, а к июню уже становятся половозрелыми медузами.
Большинство сцифомедуз раздельнополы. У медузы имеется восемь гастродермальных гонад, по одной гонаде на обеих поверхностях каждой из четырёх септ. Как правило, гаметы вымётываются через рот, но некоторые медузы вынашивают яйца на поверхности тела. Из зигот развиваются личинки планулы. После недолгого периода свободного плавания планулы оседают на субстрат, прикрепляясь к нему передним концом, претерпевают метаморфоз и превращаются в полипов.
Список источников
- Зоология беспозвоночных/под ред. В. Вестхайде и Р. Ригера. — М.: Т-во научных изданий КМК, 2008.
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Протисты и низшие многоклеточные // Зоология беспозвоночных. Функциональные и эволюционные аспекты = Invertebrate Zoology: A Functional Evolutionary Approach / пер.
 с англ. Т. А. Ганф, Н. В. Ленцман, Е. В. Сабанеевой; под ред. А. А. Добровольского и А. И. Грановича. — 7-е издание. — М.: Академия, 2008. — Т. 1. — 496 с. — 3000 экз. — ISBN 978-5-7695-3493-5
с англ. Т. А. Ганф, Н. В. Ленцман, Е. В. Сабанеевой; под ред. А. А. Добровольского и А. И. Грановича. — 7-е издание. — М.: Академия, 2008. — Т. 1. — 496 с. — 3000 экз. — ISBN 978-5-7695-3493-5
 с англ. Т. А. Ганф, Н. В. Ленцман, Е. В. Сабанеевой; под ред. А. А. Добровольского и А. И. Грановича. — 7-е издание. — М.: Академия, 2008. — Т. 1. — 496 с. — 3000 экз. — ISBN 978-5-7695-3493-5
с англ. Т. А. Ганф, Н. В. Ленцман, Е. В. Сабанеевой; под ред. А. А. Добровольского и А. И. Грановича. — 7-е издание. — М.: Академия, 2008. — Т. 1. — 496 с. — 3000 экз. — ISBN 978-5-7695-3493-5Двусторонняя симметрия кораллов древнее радиальной • Сергей Ястребов • Новости науки на «Элементах» • Эволюция, Систематика, Зоология
Коралловые полипы (Anthozoa) — древнейшая эволюционная ветвь группы стрекающих (Cnidaria), которая, в свою очередь, является сестринской по отношению к двусторонне-симметричным животным (Bilateria). В учебниках зоологии долгое время преобладало мнение, что все стрекающие — радиально-симметричные животные и что двусторонняя симметрия билатерий — уникальное для этой группы производное состояние. На самом деле у коралловых полипов внутренняя симметрия тела двусторонняя. Это вполне может оказаться примитивным состоянием и для билатерий, и для стрекающих.
Древо животного мира
Современная зоология выделяет пять главных эволюционных ветвей многоклеточных животных: губки, гребневики, пластинчатые, стрекающие (книдарии) и двусторонне-симметричные (билатерии). Подавляющее большинство животных, с которыми мы встречаемся в обычной жизни, относится к группе билатерий. Ее бесчисленные представители, от шмеля до морской звезды и от плоского червя до человека, своим разнообразием намного превосходят весь остальной животный мир вместе взятый. Билатерии — безусловно, самая эволюционно успешная группа не только среди животных, но и среди всех многоклеточных организмов на Земле. Неудивительно, что вопрос о том, как появились первые билатерии, почти полтора века интриговал зоологов, вызывая среди них оживленные споры (см., например: В. В. Малахов, 2004. Новый взгляд на происхождение билатерий).
Подавляющее большинство животных, с которыми мы встречаемся в обычной жизни, относится к группе билатерий. Ее бесчисленные представители, от шмеля до морской звезды и от плоского червя до человека, своим разнообразием намного превосходят весь остальной животный мир вместе взятый. Билатерии — безусловно, самая эволюционно успешная группа не только среди животных, но и среди всех многоклеточных организмов на Земле. Неудивительно, что вопрос о том, как появились первые билатерии, почти полтора века интриговал зоологов, вызывая среди них оживленные споры (см., например: В. В. Малахов, 2004. Новый взгляд на происхождение билатерий).
Надо признать, что проблема происхождения билатерий еще не решена. Более того, регулярно случаются новые открытия, заставляющие ученых вновь и вновь пересматривать взгляды на нее (см., например: Ксенотурбеллиды оказались близки к предкам двусторонне-симметричных животных, «Элементы», 15.02.2016). Эта тема — одна из самых актуальных в современной эволюционной биологии, по крайней мере в той ее части, которая интересуется планами строения и жизненными формами животных.
Большинство биологов традиционно полагало, что двусторонне-симметричные животные произошли от кишечнополостных — организмов, у которых симметрия тела радиальная (Radiata). Именно в соответствии с этой схемой построен, например, школьный учебник зоологии. По умолчанию считается, что радиальная симметрия примитивна, а двусторонняя производна. Вероятно, двусторонняя симметрия возникла в момент, когда кишечнополостное, обладавшее радиальной симметрией (как медуза), перешло к активному ползанью по морскому дну.
Слабость этой идеи в том, что ни радиально-симметричные животные (Radiata), ни кишечнополостные (Coelenterata), скорее всего, не являются реальными эволюционными ветвями. Это сборные группы. Под названием «кишечнополостные» раньше объединяли гребневиков и стрекающих (к последним относятся медузы, кораллы, гидры и прочие полипы). На самом деле сходство гребневиков со стрекающими — поверхностное, а родство в лучшем случае очень далекое (см., например: Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18. 09.2015). Никакого отношения к происхождению билатерий гребневики, скорее всего, не имеют.
09.2015). Никакого отношения к происхождению билатерий гребневики, скорее всего, не имеют.
Наоборот, стрекающие (Cnidaria) к билатериям очень близки. По современным данным, это их ближайшие родственники. Казалось бы, такое родство только подтверждает гипотезу о происхождении билатерий от радиально-симметричных организмов. Каждый, кто видел медузу или гидру, подтвердит, что их симметрия — радиальная, то есть лучевая. Почему бы билатериям не иметь таких предков?
Но и тут не все так просто.
Загадка кораллов
Мы уже сказали, что стрекающие и билатерии — ближайшие родственники. Иными словами, они равноправные потомки некоего предка, общего только для этих двух групп. Такие группы называются сестринскими. Очевидно, что быть предками или потомками друг друга сестринские группы не могут по определению. И в самом деле, стрекающие и билатерии появляются в палеонтологической летописи приблизительно одновременно — в конце вендского периода (см. A. Y. Ivantsov, M. A. Fedonkin, 2002. Conulariid–like fossil from the Vendian of Russia: a metazoan clade across the Proterozoic/Palaeozoic boundary). Считать, что стрекающие примитивнее билатерий, нет никаких оснований, ровно так же, как нет никаких оснований и для обратного.
Conulariid–like fossil from the Vendian of Russia: a metazoan clade across the Proterozoic/Palaeozoic boundary). Считать, что стрекающие примитивнее билатерий, нет никаких оснований, ровно так же, как нет никаких оснований и для обратного.
Но как же был устроен общий предок билатерий и стрекающих? Палеонтология ответить на этот вопрос пока не может. Значит, дело за сравнительным исследованием современных животных. Что могут нам «сообщить» стрекающие в свете сравнения с билатериями?
Стрекающие — крупная группа животных, в состав которой входит больше 10 000 современных видов. Они бывают очень разными. Крохотная пресноводная гидра совсем не похожа на роскошных медуз или на невзрачных строителей громадных коралловых рифов. Тем не менее можно утверждать, что все стрекающие делятся всего лишь на два эволюционных ствола (рис. 1). Ствол Anthozoa — это коралловые полипы, донные животные, никогда не имеющие в своем развитии стадии медузы. Ствол Medusozoa состоит из животных, у которых стадии медузы и полипа, как правило, чередуются в одном и том же жизненном цикле.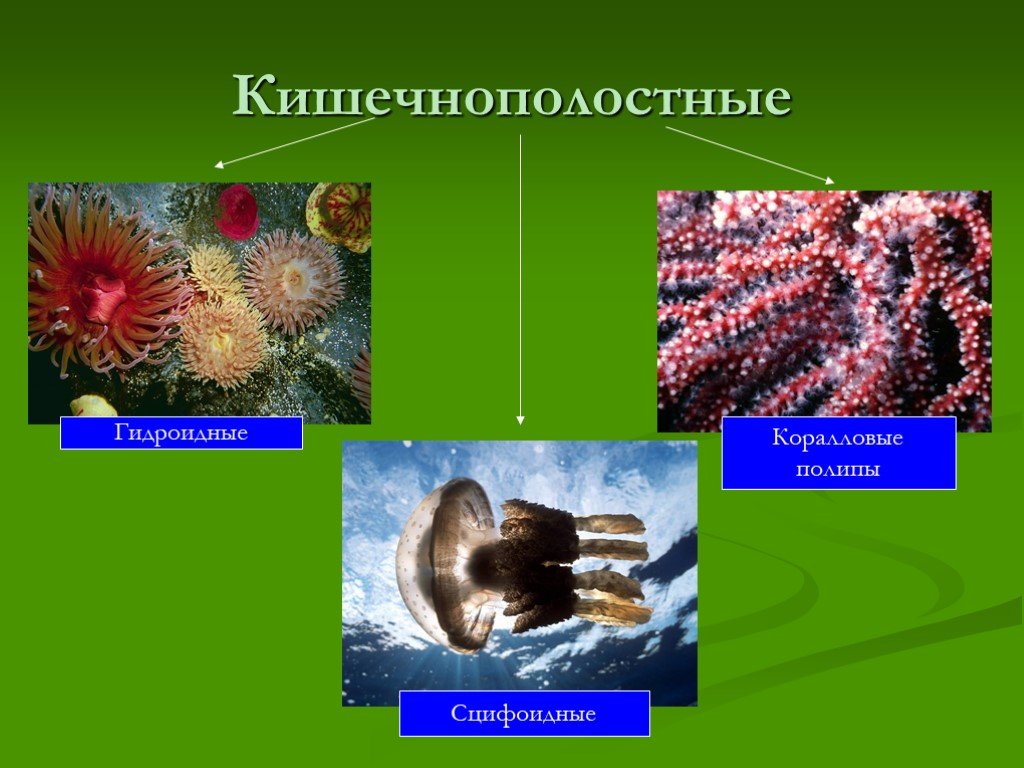 У некоторых Medusozoa или стадия медузы, или стадия полипа могут отсутствовать, но это — случаи вторичной утраты. Судя по всему, «изобретение» жизненной формы медузы и соответствующего типа жизненного цикла было важнейшим в истории стрекающих эволюционным новшеством. А положение ствола Anthozoa на эволюционном древе позволяет думать, что именно он сохранил самые древние признаки. Более того, есть работы, где предполагается, что Anthozoa — вовсе не ствол, а эволюционный уровень, охватывающий всех древнейших стрекающих (E. Kayal et al., 2013. Cnidarian phylogenetic relationships as revealed by mitogenomics). Так или иначе, примитивность этих животных несомненна.
У некоторых Medusozoa или стадия медузы, или стадия полипа могут отсутствовать, но это — случаи вторичной утраты. Судя по всему, «изобретение» жизненной формы медузы и соответствующего типа жизненного цикла было важнейшим в истории стрекающих эволюционным новшеством. А положение ствола Anthozoa на эволюционном древе позволяет думать, что именно он сохранил самые древние признаки. Более того, есть работы, где предполагается, что Anthozoa — вовсе не ствол, а эволюционный уровень, охватывающий всех древнейших стрекающих (E. Kayal et al., 2013. Cnidarian phylogenetic relationships as revealed by mitogenomics). Так или иначе, примитивность этих животных несомненна.
Коралловые полипы сами по себе довольно разнообразны (число их видов превосходит число видов всех Medusozoa, вместе взятых). Они бывают одиночными или колониальными, бывают прикрепленными или подвижными, как, например, популярная у биологов развития хищная актиния нематостелла. Тем не менее у всех коралловых полипов можно выделить как минимум две общие особенности, важные для нашей темы (рис. 2).
2).
Во-первых, перед входом в кишечную полость кораллового полипа всегда находится трубчатая глотка, отчетливо сплющенная в одной плоскости. Просвет такой глотки представляет собой более или менее широкую щель. У многих видов щелевидным становится и рот — это видно снаружи.
Во-вторых, кишечная полость кораллового полипа разделена врастающими со стороны стенок перегородками — септами — на несколько частично замкнутых камер. Точное число камер соответствует числу щупалец, окружающих рот. Некоторые авторы считали эти камеры предшественниками целомических полостей двусторонне-симметричных животных (они действительно образуются похожим способом).
Наш известный зоолог Владимир Васильевич Малахов в свежем обзоре обращает внимание на то, что внутренняя симметрия тела коралловых полипов не является радиальной. На самом деле их симметрия — двусторонняя. Особенно хорошо это заметно у одиночных шестилучевых кораллов из группы цериантарий. Щелевидный рот цериантарий ведет в сплющенную глотку, плоскость которой служит плоскостью двусторонней симметрии всего животного. На одном узком конце глотки находится сифоноглиф — продольный желобок, выстланный клетками с ресничками и предназначенный для тока воды. Вблизи другого узкого конца глотки расположена зона роста, в которой образуются новые септы. Сторону полипа, где у цериантарий находится сифоноглиф, по традиции называют «спинной», а сторону, где находится зона роста, «брюшной». Это полноценная двусторонняя симметрия — как у билатерий.
На одном узком конце глотки находится сифоноглиф — продольный желобок, выстланный клетками с ресничками и предназначенный для тока воды. Вблизи другого узкого конца глотки расположена зона роста, в которой образуются новые септы. Сторону полипа, где у цериантарий находится сифоноглиф, по традиции называют «спинной», а сторону, где находится зона роста, «брюшной». Это полноценная двусторонняя симметрия — как у билатерий.
В других группах коралловых полипов картина усложняется (например, может появляться второй сифоноглиф), но внутренняя двусторонняя симметрия, определяемая плоскостью глотки, сохраняется у них всегда. По утверждению Малахова, она есть абсолютно у всех представителей Anthozoa. А вот у Medusuzoa ее, как правило, не бывает, там симметрия чисто радиальная.
Какая симметрия первична?
Итак, самая древняя эволюционная ветвь стрекающих — это коралловые полипы, у которых нет медузы и есть внутренняя двусторонняя симметрия. По мнению Малахова, есть все основания считать, что это состояние примитивно для стрекающих в целом. Он пишет:
Он пишет:
«Билатеральную симметрию кораллов нельзя считать вторичной, развившейся под влиянием образа жизни, поскольку все Anthozoa — сидячие животные, что, как известно, способствует развитию радиальной симметрии. Билатеральную симметрию кораллов можно рассматривать как первичную, унаследованную от билатеральных предков Cnidaria, тогда как неполная радиальная симметрия, видимо, развилась под влиянием прикрепленного образа жизни.»
Биология развития подтверждает, что двусторонняя симметрия «впечатана» в структуру организма коралловых полипов очень глубоко. Малахов приводит список из полутора десятков важных регуляторных генов, активность которых четко привязана или к «спинной», или к «брюшной» стороне кораллового полипа (в основном это изучали на примере уже упоминавшейся нематостеллы). Примерно так же обстоит дело и у билатерий, хотя взаимное расположение областей активности генов у них не всегда совпадает с картинкой, которая вырисовывается у кораллов.
Получается, что радиальная симметрия стрекающих — это, скорее всего, эволюционное новшество. План строения медузы можно считать наследием древнего эволюционного прошлого не в большей степени, чем план строения морской звезды или морской лилии — несомненных представителей билатерий, освоивших когда-то радиальную симметрию из-за специфического образа жизни (см., например: Уникальность плана строения иглокожих связана с перестановкой Hox-генов, «Элементы», 18.08.2014). Двусторонняя симметрия была исходной и для билатерий, и для стрекающих. Тогда логично предположить, что общий предок этих двух групп тоже был двусторонне-симметричным, а билатерии и кораллы просто унаследовали его состояние. Первичную радиальную симметрию, которой никогда не предшествовала двусторонняя, можно найти только у одной группы животных, а именно у гребневиков. По крайней мере, таково мнение профессора Малахова.
План строения медузы можно считать наследием древнего эволюционного прошлого не в большей степени, чем план строения морской звезды или морской лилии — несомненных представителей билатерий, освоивших когда-то радиальную симметрию из-за специфического образа жизни (см., например: Уникальность плана строения иглокожих связана с перестановкой Hox-генов, «Элементы», 18.08.2014). Двусторонняя симметрия была исходной и для билатерий, и для стрекающих. Тогда логично предположить, что общий предок этих двух групп тоже был двусторонне-симметричным, а билатерии и кораллы просто унаследовали его состояние. Первичную радиальную симметрию, которой никогда не предшествовала двусторонняя, можно найти только у одной группы животных, а именно у гребневиков. По крайней мере, таково мнение профессора Малахова.
Допустим, что он прав. Какие выводы, касающиеся путей эволюции животного мира, мы можем отсюда сделать?
Билатерогастрея
Самой популярной теорией происхождения многоклеточных животных всегда была теория гастреи, предложенная в 1866 году Эрнстом Геккелем (Ernst Heinrich Philipp August Haeckel). Согласно этой теории, общий предок животных был пелагическим, то есть постоянно плавающим в толще воды, шарообразным существом, которое двигалось с помощью биения жгутиков. Геккель назвал этот гипотетический организм бластеей. Потом одно из полушарий бластеи погрузилось внутрь, и возникла гастрея — организм в форме двухслойного мешка, внутренняя полость которого стала кишечной полостью животных. Стрекающие и гребневики очень похожи по своему плану строения на геккелевскую гастрею — разумеется, слегка усложненную, как и любое реальное живое существо в сравнении с отвлеченной схемой. Впрочем, план строения билатерий тоже можно вывести из гастреи — например, в рамках так называемой архицеломатной гипотезы (см. A. Masterman, 1899. On the theory of archimeric segmentation and its bearing upon the phyletic classification of the Coelomata).
Согласно этой теории, общий предок животных был пелагическим, то есть постоянно плавающим в толще воды, шарообразным существом, которое двигалось с помощью биения жгутиков. Геккель назвал этот гипотетический организм бластеей. Потом одно из полушарий бластеи погрузилось внутрь, и возникла гастрея — организм в форме двухслойного мешка, внутренняя полость которого стала кишечной полостью животных. Стрекающие и гребневики очень похожи по своему плану строения на геккелевскую гастрею — разумеется, слегка усложненную, как и любое реальное живое существо в сравнении с отвлеченной схемой. Впрочем, план строения билатерий тоже можно вывести из гастреи — например, в рамках так называемой архицеломатной гипотезы (см. A. Masterman, 1899. On the theory of archimeric segmentation and its bearing upon the phyletic classification of the Coelomata).
Теория гастреи содержит два достаточно сильных утверждения. Из нее следует, что общий предок животных был, во-первых, пелагическим (а не донным) и, во-вторых, радиально-симметричным (а не двусторонне-симметричным). Строго говоря, ни тот, ни другой тезис не является настолько очевидным, чтобы принимать его без обсуждения. Современные палеонтологические данные свидетельствуют скорее о том, что начальные этапы эволюции животных прошли полностью на дне (А. Ю. Журавлев, 2014. Ранняя история Metazoa — взгляд палеонтолога). Тогда получается, что переход к свободному плаванию в толще воды был своего рода эволюционной авантюрой, почти как выход на сушу. Неудивительно, что он совершался несколькими группами животных независимо друг от друга и в разное время. Косвенно это подтверждается тем, что среди современных простейших родственников многоклеточных животных, которые могли бы сохранить некоторые черты их предков, пелагических форм толком нет (см. Ядра мезомицетозоев делятся синхронно, как у зародышей животных, «Элементы», 05.06.2013). «Изобретение» медузы продвинутыми, а отнюдь не примитивными стрекающими тоже хорошо вписывается в эту версию.
Строго говоря, ни тот, ни другой тезис не является настолько очевидным, чтобы принимать его без обсуждения. Современные палеонтологические данные свидетельствуют скорее о том, что начальные этапы эволюции животных прошли полностью на дне (А. Ю. Журавлев, 2014. Ранняя история Metazoa — взгляд палеонтолога). Тогда получается, что переход к свободному плаванию в толще воды был своего рода эволюционной авантюрой, почти как выход на сушу. Неудивительно, что он совершался несколькими группами животных независимо друг от друга и в разное время. Косвенно это подтверждается тем, что среди современных простейших родственников многоклеточных животных, которые могли бы сохранить некоторые черты их предков, пелагических форм толком нет (см. Ядра мезомицетозоев делятся синхронно, как у зародышей животных, «Элементы», 05.06.2013). «Изобретение» медузы продвинутыми, а отнюдь не примитивными стрекающими тоже хорошо вписывается в эту версию.
А можно ли отредактировать теорию гастреи так, чтобы она перестала противоречить «донному» сценарию эволюции животных? Да, можно. Именно это сделал в середине XX века известный шведский зоолог Карл Густав Егерстен (Karl Gustav Magnus Jagersten). Предложенная им концепция известна как теория билатерогастреи (рис. 3).
Именно это сделал в середине XX века известный шведский зоолог Карл Густав Егерстен (Karl Gustav Magnus Jagersten). Предложенная им концепция известна как теория билатерогастреи (рис. 3).
Егерстен признавал часть положений теории Геккеля, но считал, что уже бластея была не пелагическим, а придонным существом. В результате постоянного передвижения по дну у нее дифференцировались стороны тела — обращенная к грунту (брюшная) и обращенная к водной толще (спинная), — а также передний конец, на котором сосредоточились чувствительные элементы. Таким образом, бластея стала двусторонне-симметричной. Егерстен называет ее билатеробластеей.
Билатеробастея захватывала со дна пищевые частицы, обволакивая их своей брюшной стороной, на которой при каждом захвате пищи возникало временное впячивание. Постепенно это впячивание обрело устойчивость и стало постоянным. Так образовался примитивный кишечник, а билатеробластея тем самым превратилась в билатерогастрею.
Кишечник билатерогастреи открывался наружу единственным отверстием, которое было одновременно и входным (ртом), и выходным (анусом). У стрекающих такая ситуация сохранилась до сих пор. У билатерий же отверстие, ведущее в кишку, сначала сильно вытянулось вдоль оси тела, а потом закрылось посредине, разделившись тем самым надвое — на рот и анус, находящиеся, соответственно, впереди и сзади. Образование рта и ануса путем разделения единого щелевидного отверстия есть в развитии многих билатерий, и это часто рассматривают как след соответствующего этапа их эволюции (см. Вторичноротость может оказаться первичной для двусторонне-симметричных животных, «Элементы», 06.09.2015).
У стрекающих такая ситуация сохранилась до сих пор. У билатерий же отверстие, ведущее в кишку, сначала сильно вытянулось вдоль оси тела, а потом закрылось посредине, разделившись тем самым надвое — на рот и анус, находящиеся, соответственно, впереди и сзади. Образование рта и ануса путем разделения единого щелевидного отверстия есть в развитии многих билатерий, и это часто рассматривают как след соответствующего этапа их эволюции (см. Вторичноротость может оказаться первичной для двусторонне-симметричных животных, «Элементы», 06.09.2015).
Кроме того, кишечник билатерогастреи предположительно образовал несколько боковых выростов. Егерстен считал, что их было три пары. У коралловых полипов перегородки между этими выростами стали септами. А у билатерий выросты первичной кишки полностью отшнуровались от нее и разместились по бокам в виде самостоятельных мешков — целомических полостей. Подобная версия происхождения целома предлагалась несколькими авторами еще в XIX веке, Егерстен просто ее дополнил.
От билатерогастреи Егерстена вполне могли бы произойти, с одной стороны, билатерии, а с другой — коралловые полипы (рис. 4). Более того, Егерстен писал, что именно особенности анатомии коралловых полипов ему эту теорию и подсказали. Широкой популярности его взгляды не получили, потому что в них входило несколько очень спорных на тот момент утверждений. Любой сторонник теории билатерогастреи должен был признать, во-первых, что кораллы — самые примитивные стрекающие, во-вторых — что у предков стрекающих была двусторонняя симметрия, и в-третьих — что общий предок стрекающих и билатерий эволюционировал на дне. Но сейчас эти тезисы, причем все три, выглядят гораздо более похожими на правду, чем полвека назад! Проницательность Густава Егерстена, признаться, поражает. Ну а если верны отдельные составные части теории, то почему не может быть верной вся теория целиком?
Тут придется добавить ложку дегтя: предположения, с которыми согласиться невозможно, у Егерстена тоже встречаются. Например, он считал, что билатеробластее предшествовала стадия радиально-симметричной пелагической бластеи, которая плавала в толще воды и питалась путем фотосинтеза (!). Увы, современные знания делают эту версию совершенно невероятной. У животных не могло быть фотосинтезирующих предков.
Увы, современные знания делают эту версию совершенно невероятной. У животных не могло быть фотосинтезирующих предков.
Нерешенные вопросы
Владимир Васильевич Малахов, судя по его более ранним работам, считает, что стрекающие и билатерии произошли от пелагического радиально-симметричного общего предка (В. В. Малахов, 2004. Новый взгляд на происхождение билатерий). Будучи изображенным, этот предок очень похож на классическую геккелевскую гастрею. Но стоит мысленно «перенести» образование кишечника у этого существа на стадию, когда оно уже жило на дне, и теория гастреи тут же превращается в теорию билатерогастреи, которая нисколько не хуже согласуется с данными по кораллам. Разница в том, что в этом случае пелагический предок животных (если он вообще был) выглядит намного более простым.
Впрочем, даже если все наши допущения верны, это не закроет тему, а породит сразу несколько новых проблем, требующих решения. Перечислим их.
Во-первых, теория билатерогастреи неплохо объясняет происхождение стрекающих и билатерий, но очень неубедительно — происхождение губок и гребневиков. Вопрос о том, как мог выглядеть общий предок всех этих четырех групп животных, в любом случае открыт. Ни для губок, ни для гребневиков, ни для пластинчатых нет никаких серьезных свидетельств того, что они хоть когда-то имели двустороннюю симметрию. Способ их происхождения — наверняка иной.
Вопрос о том, как мог выглядеть общий предок всех этих четырех групп животных, в любом случае открыт. Ни для губок, ни для гребневиков, ни для пластинчатых нет никаких серьезных свидетельств того, что они хоть когда-то имели двустороннюю симметрию. Способ их происхождения — наверняка иной.
Во-вторых, непонятно, соответствуют ли «спинная» и «брюшная» стороны кораллового полипа спинной и брюшной сторонам билатерий. Между тем это очень важный вопрос. Ответить на него, скорее всего, сможет генетика развития — гены, регулирующие становление сторон тела у кораллов, предоставляют для этого вполне достаточно материала. В целом гены, управляющие развитием, у стрекающих примерно те же, что и у билатерий, но вот их взаимодействие может отличаться. На сходной элементной базе здесь собрана существенно другая конструкция. Какая ее часть унаследована от общего предка, предстоит еще разбираться.
В-третьих, эволюция плана строения внутри группы Anthozoa сама по себе достаточно сложна. Если у цериантарий единственный сифоноглиф находится на «спинной» стороне, то у восьмилучевых и некоторых других кораллов — на «брюшной» (рис. 2). Кроме того, многие шестилучевые кораллы имеют два сифоноглифа на противоположных сторонах глотки — в этом случае двусторонняя (билатеральная) симметрия начинает трансформироваться в двулучевую (бирадиальную). Какое из всех этих состояний самое древнее, с ходу и не скажешь.
2). Кроме того, многие шестилучевые кораллы имеют два сифоноглифа на противоположных сторонах глотки — в этом случае двусторонняя (билатеральная) симметрия начинает трансформироваться в двулучевую (бирадиальную). Какое из всех этих состояний самое древнее, с ходу и не скажешь.
В-четвертых, самые примитивные современные билатерии — ксенацеломорфы — оказались устроены гораздо проще, чем можно было бы ожидать, исходя из теории билатерогастреи (см. Ксенотурбеллиды оказались близки к предкам двусторонне-симметричных животных, «Элементы», 15.02.2016). У них нет ни сквозного кишечника, ни целомических полостей. Это — серьезное противоречие, от которого так просто не отмахнешься. Можно, конечно, предполагать, что ксенацеломорфы упростились вторично, но это требует независимых доказательств, которых пока нет.
Источник: В. В. Малахов. Симметрия и щупальцевый аппарат книдарий // Биология моря. 2016. Т. 42. № 4. С. 249–259.
Сергей Ястребов
Трехмерная оценка двусторонней симметрии ладьевидной кости: анатомическое исследование
1. Мегерле К., Харенберг П.С., Германн Г., Хельмих С. Морфология ладьевидной кости и клинические результаты реконструкции ладьевидной кости. Травма . 2012;43(3):306–310. doi: 10.1016/j.injury.2011.08.015. [PubMed] [CrossRef] [Google Scholar]
Мегерле К., Харенберг П.С., Германн Г., Хельмих С. Морфология ладьевидной кости и клинические результаты реконструкции ладьевидной кости. Травма . 2012;43(3):306–310. doi: 10.1016/j.injury.2011.08.015. [PubMed] [CrossRef] [Google Scholar]
2. Schweizer A., Fürnstahl P., Nagy L. Трехмерный компьютерный томографический анализ 11 несращений талии ладьевидной кости. Журнал хирургии кисти . 2012;37(6):1151–1158. doi: 10.1016/j.jhsa.2012.02.020. [PubMed] [CrossRef] [Google Scholar]
3. Bain G. I., Bennett J. D., MacDermid J. C., Slethaug G. P., Richards R. S., Roth J. H. Измерение деформации ладьевидного горба с помощью продольной компьютерной томографии: внутри- и межнаблюдательная вариабельность с использованием различных измерений методы. Журнал хирургии кисти . 1998;23(1):76–81. [PubMed] [Google Scholar]
4. Бхат М., Маккарти М., Дэвис Т. Р. К., Они Дж. А., Доусон С. МРТ и простая рентгенография в оценке переломов со смещением талии ладьевидной кости запястья. Журнал хирургии костей и суставов — британский том . 2004;86(5):705–713. doi: 10.1302/0301-620x.86b5.14374. [PubMed] [CrossRef] [Google Scholar]
Журнал хирургии костей и суставов — британский том . 2004;86(5):705–713. doi: 10.1302/0301-620x.86b5.14374. [PubMed] [CrossRef] [Google Scholar]
5. Buijze G. A., Jørgsholm P., Thomsen N. O. B., Björkman A., Besjakov J., Ring D. Диагностическая эффективность рентгенограмм и компьютерной томографии при смещении и нестабильности острой ладьевидной талии переломы. Журнал хирургии костей и суставов, серия A . 2012; 94(21):1967–1974. doi: 10.2106/jbjs.k.00993. [PubMed] [CrossRef] [Академия Google]
6. Форвард Д.П., Сингг Х.П., Доусон С., Дэвис Т.Р.К. Клинические результаты неправильного сращения перелома ладьевидной кости через 1 год. Журнал хирургии кисти: Европейский том . 2009;34(1):40–46. doi: 10.1177/1753193408093327. [PubMed] [CrossRef] [Google Scholar]
7. Ring D., Patterson J.D., Levitz S., Wang C., Jupiter J.B. И плоскость сканирования, и наблюдатель влияют на измерения деформации ладьевидной кости. Журнал хирургии кисти . 2005;30(4):696–701. doi: 10.1016/j.jhsa.2005.03.001. [PubMed] [CrossRef] [Академия Google]
2005;30(4):696–701. doi: 10.1016/j.jhsa.2005.03.001. [PubMed] [CrossRef] [Академия Google]
8. Ока К., Моритомо Х., Мурасе Т., Гото А., Сугамото К., Йошикава Х. Паттерны деформации запястья при несращении ладьевидной кости: трехмерный и количественный анализ. Журнал хирургии кисти . 2005;30(6):1136–1144. doi: 10.1016/j.jhsa.2005.08.004. [PubMed] [CrossRef] [Google Scholar]
9. Ока К., Мурасе Т., Моритомо Х., Гото А., Сугамото К., Йошикава Х. Паттерны костного дефекта при несращении ладьевидной кости: трехмерное и количественный анализ. Журнал хирургии кисти . 2005;30(2):359–365. doi: 10.1016/j.jhsa.2004.10.004. [PubMed] [CrossRef] [Google Scholar]
10. Врумен Дж. К., Доббе Дж. Г. Г., Йонгес Р., Стрэки С. Д., Стрикстра Г. Дж. Трехмерная оценка двусторонней симметрии лучевой и локтевой костей для планирования корректирующих операций. Журнал хирургии кисти . 2012;37(5):982–988. doi: 10.1016/j.jhsa.2011.12.035. [PubMed] [CrossRef] [Google Scholar]
11. Коберн Дж. К., Упал М. А., Криско Дж. Дж. Системы координат костей запястья. Журнал биомеханики . 2007;40(1):203–209. doi: 10.1016/j.jbiomech.2005.11.015. [PubMed] [CrossRef] [Google Scholar]
Коберн Дж. К., Упал М. А., Криско Дж. Дж. Системы координат костей запястья. Журнал биомеханики . 2007;40(1):203–209. doi: 10.1016/j.jbiomech.2005.11.015. [PubMed] [CrossRef] [Google Scholar]
12. Geissler W.B., Adams J.E., Bindra R.R., Lanzinger W.D., Slutsky D.J. Переломы ладьевидной кости: что актуально, что нет. Журнал хирургии костей и суставов — американский том . 2012;94(2):169–181. doi: 10.2106/jbjs.942icl. [PubMed] [CrossRef] [Google Scholar]
13. Dobbe J.G.G., Strackee S.D., Schreurs A.W., et al. Компьютерное планирование и навигация для корректирующей остеотомии дистального отдела лучевой кости на основе предоперационной и интраоперационной визуализации. IEEE Transactions по биомедицинской инженерии . 2011;58(1):182–190. doi: 10.1109/tbme.2010.2084576. [PubMed] [CrossRef] [Google Scholar]
14. Carelsen B., Jonges R., Strackee S.D., et al. Обнаружение in vivo динамических трехмерных моделей движения в лучезапястном суставе.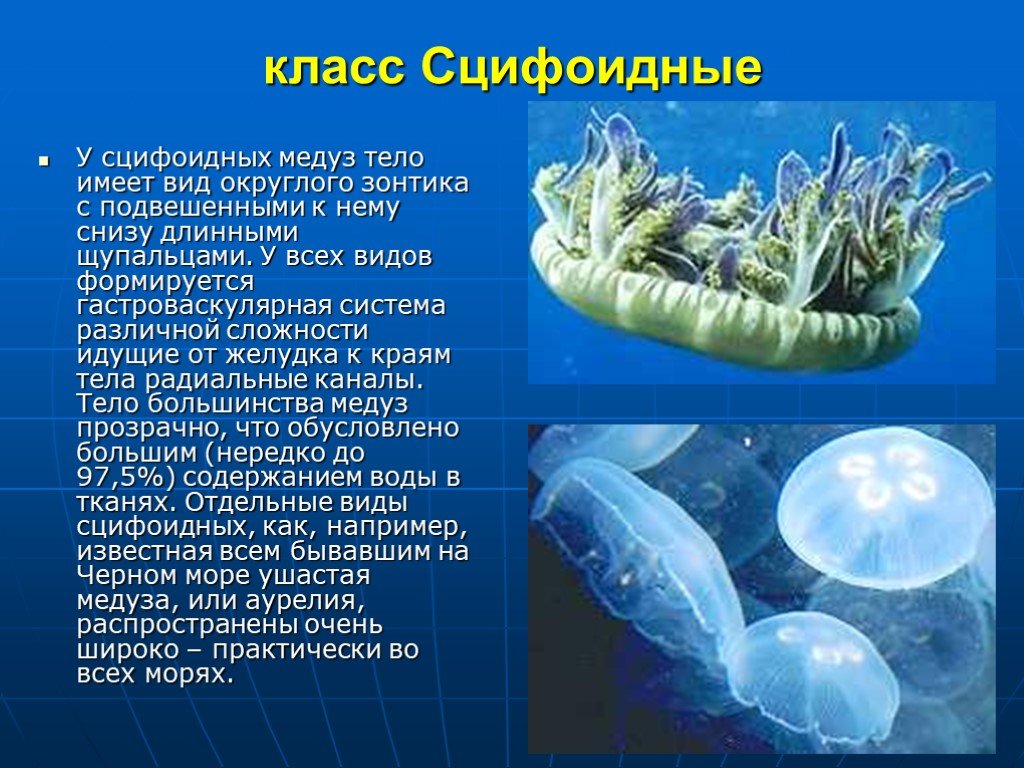 Транзакции IEEE по биомедицинской инженерии . 2009;56(4):1236–1244. doi: 10.1109/tbme.2008.2009069. [PubMed] [CrossRef] [Google Scholar]
Транзакции IEEE по биомедицинской инженерии . 2009;56(4):1236–1244. doi: 10.1109/tbme.2008.2009069. [PubMed] [CrossRef] [Google Scholar]
15. Tuijthof G. J. M., Beimers L., Jonges R., Valstar E. R., Blankevoort L. Точность метода регистрации контура кости на основе КТ для измерения относительных движений костей в заднем отделе стопы. Журнал биомеханики . 2009;42(6):686–691. doi: 10.1016/j.jbiomech.2009.01.002. [PubMed] [CrossRef] [Google Scholar]
16. Fürnstahl P., Székely G., Gerber C., Hodler J., Snedeker J.G., Harders M. Компьютерная реконструкция сложных переломов проксимального отдела плечевой кости для предоперационного планирования. Анализ медицинских изображений . 2012;16(3):704–720. doi: 10.1016/j.media.2010.07.012. [PubMed] [CrossRef] [Google Scholar]
17. Miyake J., Murase T., Oka K., Moritomo H., Sugamoto K., Yoshikawa H. Компьютерная корректирующая остеотомия при неправильно сросшихся диафизарных переломах предплечья. Журнал хирургии костей и суставов — американский том . 2012; 94, статья e150 doi: 10.2106/jbjs.k.00829. [PubMed] [CrossRef] [Google Scholar]
Журнал хирургии костей и суставов — американский том . 2012; 94, статья e150 doi: 10.2106/jbjs.k.00829. [PubMed] [CrossRef] [Google Scholar]
18. Auerbach B.M., Ruff C.B. Двусторонняя асимметрия костей конечностей: изменчивость и общность среди современных людей. Журнал эволюции человека . 2006;50(2):203–218. doi: 10.1016/j.jhevol.2005.09.004. [PubMed] [CrossRef] [Google Scholar]
19. Lazenby R. A. Скелетная биология, функциональная асимметрия и происхождение «ручности» Журнал теоретической биологии . 2002;218(1):129–138. doi: 10.1006/jtbi.2002.3052. [PubMed] [CrossRef] [Google Scholar]
20. Смит Д. К. Анатомические особенности ладьевидной кости запястья: проверка биометрических измерений и симметрии с помощью трехмерной МРТ. Радиология . 1993;187(1):187–191. doi: 10.1148/radiology.187.1.8451410. [PubMed] [CrossRef] [Google Scholar]
21. Heinzelmann A.D., Archer G., Bindra R.R. Антропометрия ладьевидной кости человека.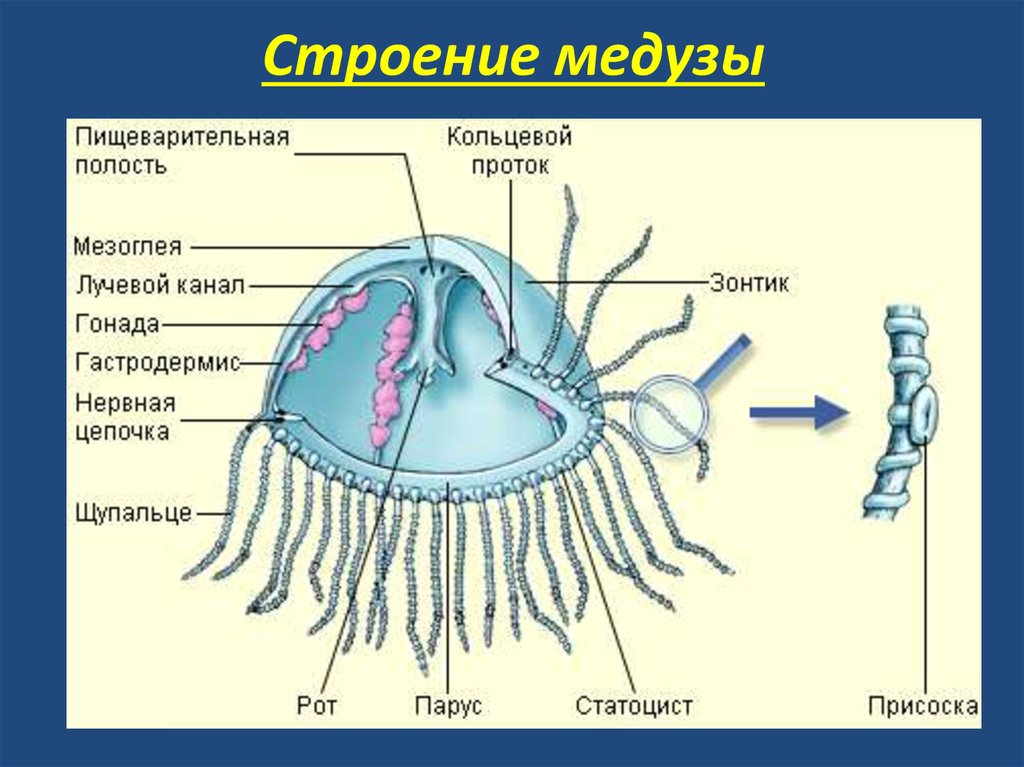 Журнал хирургии кисти . 2007;32(7):1005–1008. doi: 10.1016/j.jhsa.2007.05.030. [PubMed] [CrossRef] [Google Scholar]
Журнал хирургии кисти . 2007;32(7):1005–1008. doi: 10.1016/j.jhsa.2007.05.030. [PubMed] [CrossRef] [Google Scholar]
22. Фукуда С., Исида О., Кидо М., Судзумура Ф., Икута Ю. Морфологическое исследование ладьевидной кости с использованием математической техники и сравнительное исследование трех- размерные измерения ладьевидной кости. Хирургия кисти . 2003;8(2):157–161. [PubMed] [Google Scholar]
23. Guo Y., Tian G.L. Длина и положение длинной оси ладьевидной кости, измеренные путем анализа трехмерных реконструкций изображений компьютерной томографии. Журнал хирургии кисти: Европейский том . 2011;36(2):98–101. doi: 10.1177/1753193410377837. [PubMed] [CrossRef] [Google Scholar]
24. Letta C., Schweizer A., Fürnstahl P. Количественная оценка контралатеральных различий ладьевидной кости: сравнение геометрии кости в трех измерениях. Международный исследовательский центр анатомии . 2014;2014:5. doi: 10.1155/2014/904275.904275 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
несращение ладьевидной талии.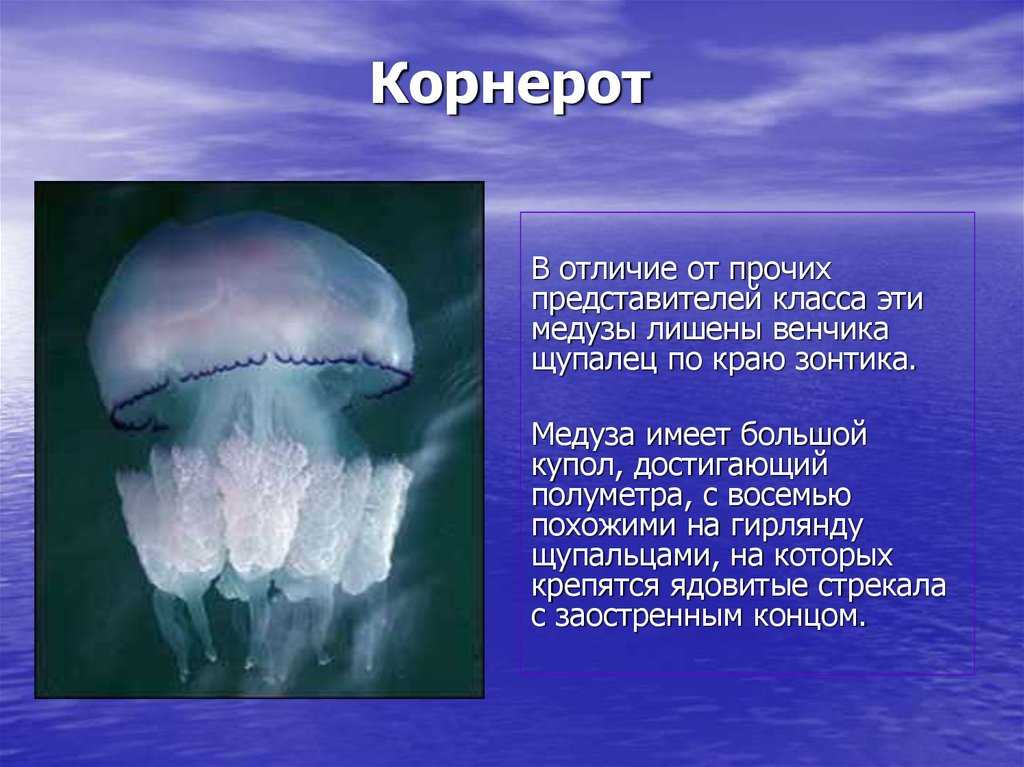 Журнал хирургии кисти . 1991;16(5):899–906. doi: 10.1016/s0363-5023(10)80158-3. [PubMed] [CrossRef] [Google Scholar]
Журнал хирургии кисти . 1991;16(5):899–906. doi: 10.1016/s0363-5023(10)80158-3. [PubMed] [CrossRef] [Google Scholar]
26. Моритомо Х., Мурасе Т., Ока К., Танака Х., Йошикава Х., Сугамото К. Взаимосвязь между местом перелома и кинематической картиной при несращении ладьевидной кости. Журнал хирургии кисти . 2008;33(9):1459–1468. doi: 10.1016/j.jhsa.2008.05.035. [PubMed] [CrossRef] [Google Scholar]
27. Амадио П. С., Берквист Т. Х., Смит Д. К., Иллструп Д. М., Куни В. П., III, Линшейд Р. Л. Неправильное сращение ладьевидной кости. Журнал хирургии кисти . 1989;14(4):679–687. doi: 10.1016/0363-5023(89)90191-3. [PubMed] [CrossRef] [Google Scholar]
28. Lynch N.M., Linscheid R.L. Корректирующая остеотомия при неправильном сращении ладьевидной кости: техника и оценка долгосрочного наблюдения. Журнал хирургии кисти . 1997;22(1):35–43. doi: 10.1016/s0363-5023(05)80177-7. [PubMed] [CrossRef] [Google Scholar]
29. Накамура Р. , Имаэда Т., Миура Т. Неправильное сращение ладьевидной кости. Журнал хирургии костей и суставов — британский том . 1991;73(1):134–137. [PubMed] [Google Scholar]
, Имаэда Т., Миура Т. Неправильное сращение ладьевидной кости. Журнал хирургии костей и суставов — британский том . 1991;73(1):134–137. [PubMed] [Google Scholar]
30. Lindstrom G., Nystrom A. Частота посттравматического артроза после первичного заживления переломов ладьевидной кости: клинико-рентгенологическое исследование. Журнал хирургии кисти . 1990;15(1):11–13. [PubMed] [Google Scholar]
31. Берджесс Р. К. Влияние имитации неправильного сращения ладьевидной кости на движение запястья. Журнал хирургии кисти . 1987; 12(5, часть 1):774–776. doi: 10.1016/s0363-5023(87)80067-9. [PubMed] [CrossRef] [Академия Google]
32. Йиранек В. А., Руби Л. К., Миллендер Л. Б., Банкофф М. С., Ньюберг А. Х. Отдаленные результаты костной пластики по Руссе: эффект неправильного сращения ладьевидной кости. Журнал хирургии костей и суставов — американский том . 1992;74(8):1217–1228. [PubMed] [Google Scholar]
Перелом ладьевидной кости — John Erickson, MD
Ладьевидная кость — одна из восьми мелких костей запястья, называемых «костями запястья». Ладьевидная кость имеет решающее значение для нормального функционирования и диапазона движений запястья, поскольку она служит важным связующим звеном между двумя рядами костей запястья.
Ладьевидная кость имеет решающее значение для нормального функционирования и диапазона движений запястья, поскольку она служит важным связующим звеном между двумя рядами костей запястья.
Из всех костей запястья ладьевидная чаще всего ломается при травмах. Перелом ладьевидной кости может произойти по разным причинам, включая спортивные травмы, падения и автомобильные столкновения. Однако наиболее частым сценарием является падение молодого человека на вытянутую руку. Обычно люди сначала не замечают травмы. Как только боль и отек от травмы уменьшатся, люди могут подумать, что травма была «просто растяжением». Часто диагноз задерживается на несколько недель или месяцев.
Каковы проблемы с переломами ладьевидной кости?
Кости живые; как и все живые ткани, они нуждаются в постоянном поступлении питательных веществ, чтобы оставаться здоровыми. Когда кровоснабжение кости уменьшается, кость может стать хрупкой и слабой. Ладьевидная кость, естественно, имеет плохое кровоснабжение по сравнению с другими костями в организме.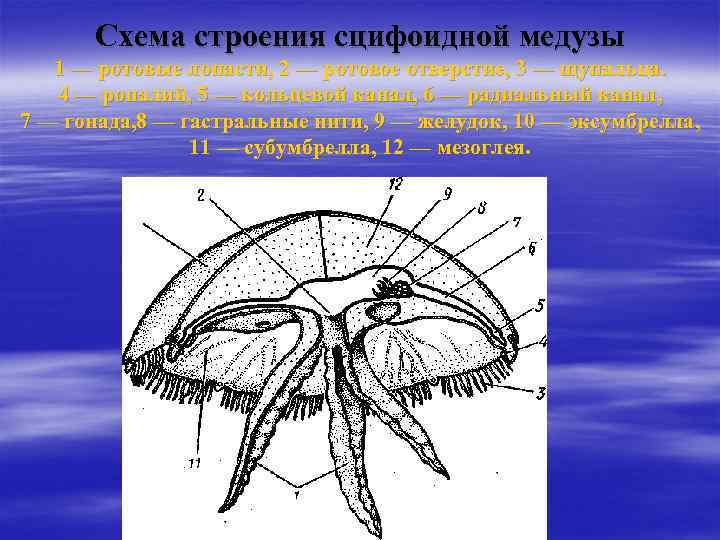 Когда ладьевидная кость ломается, кровоснабжение части кости может быть нарушено. Это подвергает переломы ладьевидной кости риску «несращения» или невозможности заживления. Кроме того, часть кости может отмирать, что известно как «аваскулярный некроз». Наконец, если перелом ладьевидной кости не заживает должным образом, со временем может развиться артрит запястья, что приведет к боли, ограничению диапазона движений запястья и снижению функции. Это состояние называется «продвинутый коллапс ладьевидной кости без сращения (SNAC)».
Когда ладьевидная кость ломается, кровоснабжение части кости может быть нарушено. Это подвергает переломы ладьевидной кости риску «несращения» или невозможности заживления. Кроме того, часть кости может отмирать, что известно как «аваскулярный некроз». Наконец, если перелом ладьевидной кости не заживает должным образом, со временем может развиться артрит запястья, что приведет к боли, ограничению диапазона движений запястья и снижению функции. Это состояние называется «продвинутый коллапс ладьевидной кости без сращения (SNAC)».
Как диагностируется перелом?
Сбор анамнеза, физикальный осмотр и несколько рентгенограмм важны для диагностики перелома ладьевидной кости. Пациенты с переломами ладьевидной кости часто болезненны при прикосновении к основанию большого пальца в области «анатомической сальниковой коробки» запястья. Во многих случаях рентгеновские снимки вскоре после травмы выглядят нормальными, и диагноз может быть отложен. Если пациенты болезненны в «табакерке», врачи часто лечат их так, как будто у них перелом ладьевидной кости, даже если первоначальные рентгенограммы в норме. Через 2-3 недели перелом становится отчетливым на рентгенограммах. И КТ, и МРТ могут быть полезны для оценки различных характеристик перелома ладьевидной кости и могут использоваться для постановки более раннего диагноза, чем рентген.
Через 2-3 недели перелом становится отчетливым на рентгенограммах. И КТ, и МРТ могут быть полезны для оценки различных характеристик перелома ладьевидной кости и могут использоваться для постановки более раннего диагноза, чем рентген.
Какие есть варианты лечения?
План лечения будет зависеть от тяжести перелома, отмеченного на рентгеновских снимках, а также от состояния здоровья пациента, уровня активности и желаний. Пациентов с правильно расположенными или «несмещенными» переломами можно лечить без хирургического вмешательства. Обычно для этого требуется гипсовая повязка, иммобилизующая запястье на 2-3 месяца. Последующие рентгеновские снимки получают в клинике, чтобы оценить, как заживает перелом. Терапия часто помогает улучшить диапазон движений и силу после заживления перелома. Иногда пациенты с переломами без смещения предпочитают хирургическое вмешательство, чтобы сократить время наложения гипсовой повязки.
У некоторых пациентов переломы со смещением, нестабильность или риск незаживления в гипсовой повязке. Таким пациентам может быть рекомендовано хирургическое вмешательство. Цель операции — стабилизировать перелом с помощью имплантата и способствовать заживлению ладьевидной кости.
Таким пациентам может быть рекомендовано хирургическое вмешательство. Цель операции — стабилизировать перелом с помощью имплантата и способствовать заживлению ладьевидной кости.
Что связано с хирургией?
Операция проводится амбулаторно, часто с применением регионарной анестезии (блокада нервов) с седацией. Во время операции я улучшаю положение перелома и использую металлический имплантат для стабилизации перелома. Интраоперационный рентген, называемый рентгеноскопией, используется для подтверждения выравнивания. В большинстве случаев для фиксации кости я использую компрессионный винт для ладьевидной кости. Это называется «открытая репозиция и внутренняя фиксация». Иногда костный трансплантат используется для улучшения выравнивания перелома и потенциала заживления перелома. Костный трансплантат можно взять из близлежащей лучевой кости или гребня подвздошной кости таза.
Чего ожидать после операции?
Ваше запястье и большой палец будут защищены шиной, которую снимут в клинике вместе с кожными швами примерно через 10-14 дней. В этот момент будет наложена гипсовая повязка или шина для защиты перелома до тех пор, пока заживление не будет отмечено на рентгенограмме или компьютерной томографии. Я рекомендую избегать поднятия тяжестей, тяги, толкания или хватания примерно в течение 3-4 месяцев после операции.
В этот момент будет наложена гипсовая повязка или шина для защиты перелома до тех пор, пока заживление не будет отмечено на рентгенограмме или компьютерной томографии. Я рекомендую избегать поднятия тяжестей, тяги, толкания или хватания примерно в течение 3-4 месяцев после операции.
Боль, отек и скованность в запястье постепенно уменьшаются со временем. Большинство пациентов могут вернуться к более нормальной функции примерно через 4-6 месяцев после операции, но максимальное улучшение может занять несколько месяцев. Артрит, также известный как дегенеративное заболевание суставов, может развиться в будущем, если первоначальная травма повредила поверхность сустава. Моя цель — помочь каждому пациенту восстановить максимально возможную функцию после травмы.
Что я могу сделать, чтобы улучшить результаты операции?
Адекватное питание играет важную роль в процессе выздоровления. Я рекомендую диету, богатую белком и кальцием, и адекватное потребление калорий в день, пока ваше тело заживает.